Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Vectoren en klonering in hogere organismen : dierlijke cellen
(Primrose & Twyman : hoofdstuk 12) Onderzoeksbelang : studie genfunctie - genregulatie aanmaak recombinante eiwitten - juiste posttranslationele modificaties - vorming van correcte disulfidebruggen protein disulfide isomerase (PDI) in endoplasmatisch reticulum van alle eukaryoten ; vorming van S-S bruggen ; eveneens chaperone. - aminozuurverwijdering van het oorspronkelijke polypeptide - O-linked of N-linked glycosylaties => ongeveer 30% van de eukaryotische proteinen zijn geglycosyleerd manipuleren van endogeen genoom => gentherapie Commercieel : synthese, o.a. vaccins, hormonen in zoogdier-celculturen “Gene delivery” voor dierlijke cellen in vivo versus in vitro : gentherapie 4 hoofdstrategiën voor horizontale gentransfer chemische - fysische “transfectie”/transformatie DNA-”delivery” met bacteriële vectoren virussen als gentranfervectoren Plasmidevectoren : dierlijke + bacteriële componenten Selectiemerkers
 Onderzoeksbelang : studie genfunctie - genregulatie. aanmaak recombinante eiwitten - juiste posttranslationele modificaties. - vorming van correcte disulfidebruggen. protein disulfide isomerase (PDI) in endoplasmatisch reticulum. van alle eukaryoten ; vorming van S-S bruggen ; eveneens chaperone. - aminozuurverwijdering van het oorspronkelijke polypeptide. - O-linked of N-linked glycosylaties. => ongeveer 30% van de eukaryotische proteinen zijn geglycosyleerd. manipuleren van endogeen genoom => gentherapie. Commercieel : synthese, o.a. vaccins, hormonen in zoogdier-celculturen. Gene delivery voor dierlijke cellen in vivo versus in vitro : gentherapie. 4 hoofdstrategiën voor horizontale gentransfer. chemische - fysische transfectie /transformatie. DNA- delivery met bacteriële vectoren. virussen als gentranfervectoren. Plasmidevectoren : dierlijke + bacteriële componenten. Selectiemerkers.")
2
* Gentransfer (DNA- transfer) naar dierlijke cellen
(DNA opname : Primrose & Twyman, pp ) 4 hoofdstrategiën van transformatie “niet-biologisch” Transformatie - fysisch Transformatie - chemisch – endocytose ‘Bactofection’ – bacterie-gemediëerd Transductie – virus-gemedieerd “biologisch” *
 4 hoofdstrategiën van transformatie. niet-biologisch Transformatie - fysisch. Transformatie - chemisch – endocytose. ‘Bactofection’ – bacterie-gemediëerd. Transductie – virus-gemedieerd. biologisch *")
3
N DNA transfer via chemische of fysische weg
Chemische methoden – transformatie (transfectie) complex zodanig dat interactie met celmembraan bevorderd wordt vb. Positieve lading opname door endocytose Lipofiel complex membraanfusie DNA in cytoplasma Fysische methoden – transformatie (transfectie) door fysische kracht introductie van ‘naakt’ DNA vb. micro-injectie – “particle bombardment” – electroporatie – ultrasone behandeling N
 complex zodanig dat interactie met celmembraan bevorderd wordt. vb. Positieve lading opname door endocytose. Lipofiel complex membraanfusie DNA in cytoplasma. Fysische methoden – transformatie (transfectie) door fysische kracht introductie van ‘naakt’ DNA. vb. micro-injectie – particle bombardment – electroporatie. – ultrasone behandeling N.")
4
N Chemische methoden – transformatie (transfectie)
Calciumfosfaat co-precipitaat methode - DNA/calciumfosfaat co-precipitaat afzetting op de membraan en daarna internalisatie door endocytose - migreert naar de nucleus voor expressie - “coating” van cellen door het co-precipitaat is nodig ; werkt daarom best voor cellen in monolayer- of suspensie-cultuur, maar niet erg geschikt voor cellen in “clumps” (klompjes) - 1-2% van de cellen nemen DNA op - slechts beperkte stabiele transformatie - optimalisatie via specifieke buffersystemen voor trage precipitatie en optimale balans tussen DNA, calciumfosfaat, buffer/pH N
 - 1-2% van de cellen nemen DNA op - slechts beperkte stabiele transformatie. - optimalisatie via specifieke buffersystemen voor trage precipitatie en. optimale balans tussen DNA, calciumfosfaat, buffer/pH. N.")
5
N Diethylaminoethyl dextraan dextraan Gebruik van polyplexen
DEAE Gebruik van polyplexen - DEAE-dextraan (diethylaminoethyldextraan) – een polykationisch polymeer koolhydraat in complex met DNA - polykationische componenten die spontaan oplosbare complexen vormen (“polyplexen”) - elektrostatische interactie met DNA - initiëel ontwikkeld voor “probleem”cellen die moeilijk te transformeren waren met de calciumfosfaat methode - nieuwe generatie polykationische componenten (zie Tabel 12.1 : Primrose & Twyman) bvb. poly-L-lysine, spermine, polyethylenimine, dendrimeren synthetische polyaminen (veel aminogroepen - positieve ladingen) - ook voor in vivo gentransfer : controle over genexpressie en ‘gene delivery’ via specifieke polymeren met veranderende eigenschappen bvb. onder invloed van temperatuursregulatie N
 – een polykationisch polymeer koolhydraat in complex met DNA. - polykationische componenten die spontaan oplosbare complexen. vormen ( polyplexen ) - elektrostatische interactie met DNA. - initiëel ontwikkeld voor probleem cellen die moeilijk te transformeren. waren met de calciumfosfaat methode. - nieuwe generatie polykationische componenten (zie Tabel 12.1 : Primrose & Twyman) bvb. poly-L-lysine, spermine, polyethylenimine, dendrimeren. synthetische polyaminen (veel aminogroepen - positieve ladingen) - ook voor in vivo gentransfer : controle over genexpressie en ‘gene delivery’ via specifieke polymeren met veranderende eigenschappen. bvb. onder invloed van temperatuursregulatie. N.")
6
N Liposomen en lipoplexen
- verpakking in liposoom (artificiële, ‘fusogene’ fosfolipidevesikels) => membraanfusie (eerste voorbeeld : Schaffner in 1980 : fusie van bacteriële protoplasten met dierlijke cellen om DNA binnen te brengen) - kationische/neutrale lipidemengsels vormen stabiele complexen met DNA => lipoplexen - mogelijkheid tot transformatie van grote DNA’s = “zachte” weg - hoge efficiëntie van transformatie (tot 90% van de cellen) – zowel transiënt als stabiel - transformatie m.b.v. liposomen = lipofectie - mogelijkheid van transformatie van cellen in levende organismen na injectie in doelwit- cellen (‘gene delivery’) : optimalisatie door insertie van virale eiwitten die celfusie promoten of targeting naar specifieke cellen. Kationisch liposoom in interactie met DNA = lipoplex N
 => membraanfusie. (eerste voorbeeld : Schaffner in 1980 : fusie van bacteriële protoplasten met. dierlijke cellen om DNA binnen te brengen) - kationische/neutrale lipidemengsels vormen stabiele complexen met DNA. => lipoplexen. - mogelijkheid tot transformatie van grote DNA’s = zachte weg. - hoge efficiëntie van transformatie (tot 90% van de cellen) – zowel transiënt. als stabiel. - transformatie m.b.v. liposomen = lipofectie. - mogelijkheid van transformatie van cellen in. levende organismen na injectie in doelwit- cellen (‘gene delivery’) : optimalisatie door. insertie van virale eiwitten die. celfusie promoten of targeting. naar specifieke cellen. Kationisch liposoom in interactie met DNA = lipoplex. N.")
7
N Fysische methoden – transformatie (transfectie) Elektroporatie
- transiënte vorming van poriën (nm-grootte) - blootstelling aan elektrische puls - intensiteit/tijd - empirische optimalisatie - eenvoudig - reproduceerbaar - mogelijkheid in vivo gentransfer - op oppervlak met elektroden/naaldelektroden Ultrasone transfectie snel oscillerende probe (tip sonicator) ‘inklappen’ van membranen - ‘cavities’ – transiënte doorgang van DNA door membranen zowel in vitro als in vivo ‘gene delivery’ Micro-injectie - directe transfer tot in de cel - enkel toepasbaar voor beperkt aantal cellen - meestal toegepast op grote celtypes, bvb. oöcyten - in vivo en in vitro “Particle bombardment” - initieel ontwikkeld voor transformatie van planten - kleine metalen (gouden) partikels worden gecoat met DNA en m.b.v. kracht geïntroduceerd N
 - blootstelling aan elektrische puls - intensiteit/tijd - empirische optimalisatie. - eenvoudig - reproduceerbaar. - mogelijkheid in vivo gentransfer - op oppervlak met elektroden/naaldelektroden. Ultrasone transfectie. snel oscillerende probe (tip sonicator) ‘inklappen’ van membranen - ‘cavities’ – transiënte doorgang van DNA door. membranen. zowel in vitro als in vivo ‘gene delivery’ Micro-injectie. - directe transfer tot in de cel. - enkel toepasbaar voor beperkt aantal cellen. - meestal toegepast op grote celtypes, bvb. oöcyten. - in vivo en in vitro. Particle bombardment - initieel ontwikkeld voor transformatie van planten. - kleine metalen (gouden) partikels worden gecoat met DNA. en m.b.v. kracht geïntroduceerd. N.")
8
N DNA transfer via biologische weg : bactofection
Bacteriële gentransfer – bacteriële vectoren Transfer naar planten, m.b.v. Agrobacterium tumefaciens (zie hoofdstuk 16 Plant) A. tumefaciens kan ook DNA introduceren in dierlijke cellen (2001) A. tumefaciens : - transfer zonder invasie (blijft in de extracellulaire ruimte) - aanhechting en vorming pilus : DNA transfer via een conjugatief mechanisme Bactofection van dierlijke cellen door invasie met de bacteriën : - o.a. Salmonella spp., Listeria monocytogenese, Shigella flexneri (mogen niet pathogeen zijn, kunnen desondanks storende neveneffecten veroorzaken bij lyse ; of reacties bij gentherapeutische toepassingen) - invasief in cytoplasma (Listeria, Shigella) of in vacuole (Salmonella, Yersinia) opname in de cel, vervolgens lyse + vrijstelling van plasmide migreert naar de nucleus ; expressie naargelang de expressiesignalen - verzwakking (‘attenuation’) nodig - zoniet vernietiging van de gastheercellen auxotrofe mutanten of stammen met induceerbare autolysis N
 A. tumefaciens kan ook DNA introduceren in dierlijke cellen (2001) A. tumefaciens : - transfer zonder invasie (blijft in de extracellulaire ruimte) - aanhechting en vorming pilus : DNA transfer via een conjugatief mechanisme. Bactofection van dierlijke cellen door invasie met de bacteriën : - o.a. Salmonella spp., Listeria monocytogenese, Shigella flexneri. (mogen niet pathogeen zijn, kunnen desondanks storende neveneffecten veroorzaken bij lyse ; of reacties bij gentherapeutische toepassingen) - invasief in cytoplasma (Listeria, Shigella) of in vacuole (Salmonella, Yersinia) opname in de cel, vervolgens lyse + vrijstelling van plasmide. migreert naar de nucleus ; expressie naargelang de expressiesignalen. - verzwakking (‘attenuation’) nodig - zoniet vernietiging van de gastheercellen. auxotrofe mutanten of stammen met induceerbare autolysis. N.")
9
Toepassingen : o. a. gentransfer in vitro en in vivo : vb
Toepassingen : o.a. gentransfer in vitro en in vivo : vb. afleveren van recombinante DNA vaccins N nb. Bij ‘alternatieve gentherapie’ worden de bacteriën actief behouden en wordt het eiwit in situ (in vivo) tot expressie gebracht door de bacterie. Vergelijking van de voornaamste vectortypes in functie van diverse parameters Virale vectoren Niet-virale vectors Bacteriële vectoren Veiligheid Efficiëntie Lage productiekost Eenvoud van productie Eenvoudige ‘delivery’ Hoeveelheid toegeleverd DNA
 tot expressie gebracht door de bacterie. Vergelijking van de voornaamste vectortypes in functie van diverse parameters. Virale vectoren Niet-virale vectors Bacteriële vectoren. Veiligheid Efficiëntie Lage productiekost Eenvoud van productie Eenvoudige ‘delivery’ Hoeveelheid toegeleverd DNA")
10
DNA transfer via biologische weg :
De eerste experimenten (met fysische en chemische transformatiemethoden) werden initiëel toegepast met complexe mengsels van DNA of afzonderlijke gensequenties, bij gebrek aan vectorsystemen. Met de karakterisatie van het SV40 genoom kwam de eerste mogelijkheid om virale en nadien plasmidevectoren te ontwikkelen Gebruik van plasmidevectoren : Voordelen : - gemak van in vitro manipulaties - introductie van modulaire elementen (e.g. promotersequenties) die onafhankelijk van het transgen bruikbaar zijn - selectiemerker op het construct omzeilt de noodzaak van co-transformatie (transgen en selectiemerker zijn gekoppeld : zie verder) - mogelijkheid tot constructie van pendelvectoren - vereisen geen integratie in het genoom - episomaal stabiel Runaway replicon : een replicon zonder, of induceerbaar tot verlies van, ’copy number’ controle : deze afwezigheid of verlies leidt tot ongecontroleerde replicatie en extreme toename van het aantal kopijen van het DNA waarin dit replicon ingebouwd is. Celdood kan een gevolg zijn zodat toepassing vooral op DNA en eiwitproductie gericht is. Zie voorbeelden verderop. Virus- (of plasmide-)gemedieerd : transductie Hier, gezien de historische context, behandeld samen met het exploreren van fysische of chemische transformatiemethoden, aangezien dit samenging met de opbouw van de eerste dierlijke virusvectoren ; transformatie met “vrij” DNA of met een replicon.
 werden initiëel toegepast met complexe mengsels van DNA of afzonderlijke gensequenties, bij gebrek aan vectorsystemen. Met de karakterisatie van het SV40 genoom kwam de eerste mogelijkheid om virale en nadien plasmidevectoren te ontwikkelen. Gebruik van plasmidevectoren : Voordelen : - gemak van in vitro manipulaties. - introductie van modulaire elementen (e.g. promotersequenties) die onafhankelijk van. het transgen bruikbaar zijn. - selectiemerker op het construct omzeilt de noodzaak van co-transformatie (transgen en. selectiemerker zijn gekoppeld : zie verder) - mogelijkheid tot constructie van pendelvectoren - vereisen geen integratie in het. genoom - episomaal stabiel. Runaway replicon : een replicon zonder, of induceerbaar tot verlies van, ’copy number’ controle : deze afwezigheid of verlies leidt tot ongecontroleerde replicatie en extreme toename van het aantal kopijen van het DNA waarin dit replicon ingebouwd is. Celdood kan een gevolg zijn zodat toepassing vooral op DNA en eiwitproductie gericht is. Zie voorbeelden verderop. Virus- (of plasmide-)gemedieerd : transductie. Hier, gezien de historische context, behandeld samen met het exploreren van fysische of chemische transformatiemethoden, aangezien dit samenging met de opbouw van de eerste dierlijke virusvectoren ; transformatie met vrij DNA of met een replicon.")
11
Transformatie gebeurt in twee ‘stadia’ :
introductie van DNA in de cel (transfectie = algemene term voor onderscheid met infectie) incorporatie in de nucleus (veelal door integratie in het gastheerchromosoom) Resultaat van transformatie : indien tijdelijke toestand = transiënte transformatie/transfectie - effect kortstondig onderscheid dus tussen transiënte versus permanente (‘stabiele’) transformatie => ‘amplificatie’ mogelijk met methotrexaat integratie in het genoom - is overerfbaar - kan leiden tot stabiele transgene cellijn integratie = inefficiënt selectie is vereist "Klonering” => het betreft hier transformatie van somatische cellen in cultuur (voor wijziging van ‘germline’ cellen en eventuele aanmaak van transgene dieren zijn er andere vereisten) ; slechts een kleine fractie van het inkomende DNA zal integreren in het cellulair genoom maar kan aanleiding geven tot een nieuwe cellijn met nieuwe karakteristiek. => principe van ‘co-transformatie’ : het binnenkomende DNA concatemeriseert tot grote moleculen vooraleer het integreert in het genoom. Twee verschillende DNA’s hoeven dus niet gekoppeld te worden (mag uiteraard wel) : dit gebeurt in de cel. Een gen samen met een selecteerbare merker bleken samen (in elkaars buurt) geïntegreerd te zijn.
 incorporatie in de nucleus (veelal door integratie in het gastheerchromosoom) Resultaat van transformatie : indien tijdelijke toestand = transiënte transformatie/transfectie - effect kortstondig. onderscheid dus tussen transiënte versus permanente (‘stabiele’) transformatie. => ‘amplificatie’ mogelijk met methotrexaat. integratie in het genoom - is overerfbaar - kan leiden tot stabiele transgene cellijn. integratie = inefficiënt selectie is vereist. Klonering => het betreft hier transformatie van somatische cellen in cultuur (voor wijziging. van ‘germline’ cellen en eventuele aanmaak van transgene dieren zijn er andere vereisten) ; slechts een kleine fractie van het inkomende DNA zal integreren in het cellulair. genoom maar kan aanleiding geven tot een nieuwe cellijn met nieuwe karakteristiek. => principe van ‘co-transformatie’ : het binnenkomende DNA concatemeriseert tot. grote moleculen vooraleer het integreert in het genoom. Twee verschillende DNA’s. hoeven dus niet gekoppeld te worden (mag uiteraard wel) : dit gebeurt in de cel. Een gen. samen met een selecteerbare merker bleken samen (in elkaars buurt) geïntegreerd te zijn.")
12
* Merkers en selectie, en reporters
Reporters : (verdere informatie in Primrose & Twyman : pp en ) - CAT : radiochemisch, chromatografie, ELISA (intracellulair) - Alkalische fosfatase : kleur, fluorografie (secretie) - LacZ : kleurvorming, ... - GusA : kleurvorming, (gusA gen = uidA gen : glucuronidase-gen) - luciferase (luciferine, ATP, Mg2+) : flash - GFP (UV/blauw => groen fluorescent) en varianten (vereist geen substraat ; vooral nuttig voor localisatie-experimenten) * Merkers : - uit het nucleotidemetabolisme (TK, HPRT, APRT, ...) - exogene, dominante (neo, nptII (aphII) : geneticine; ...) - "amplificatie" : methotrexaat : competitieve inhibitie van DHFR resistentie door - mutaties in DHFR - verhindering opname - verhoogde doelwitdosis
 - CAT : radiochemisch, chromatografie, ELISA (intracellulair) - Alkalische fosfatase : kleur, fluorografie (secretie) - LacZ : kleurvorming, GusA : kleurvorming, ... (gusA gen = uidA gen : glucuronidase-gen) - luciferase (luciferine, ATP, Mg2+) : flash. - GFP (UV/blauw => groen fluorescent) en varianten. (vereist geen substraat ; vooral nuttig voor localisatie-experimenten) * Merkers : - uit het nucleotidemetabolisme (TK, HPRT, APRT, ...) - exogene, dominante (neo, nptII (aphII) : geneticine; ...) - amplificatie : methotrexaat : competitieve inhibitie van DHFR. resistentie door - mutaties in DHFR. - verhindering opname. - verhoogde doelwitdosis.")
13
Endogene selectiemerkers - bruikbaar bij mutante cellijnen
Drie types selectiemerkers in zoogdiercellen Endogene selectiemerkers - bruikbaar bij mutante cellijnen Dominante selectiemerkers ‘Kwantitatieve’ selectiemerkers Endogene selectiemerkers (Primrose & Twyman, voor een korte lijst in Tabel 12.2 en beschrijving in Fig 12.3) Voorbeeld: Tk = thymidinekinasegen historisch : experiment met muiscellen deficiënt in TK - te transformeren met Herpes Simplex Virus (HSV) Tk gen => resultaat : activiteit herwonnen transformanten te selecteren op HAT-medium (hypoxanthine - aminopterine - thymidine) : aminopterine blokkeert de de novo synthese van dTMP (en dus van dTTP) voor achtergrond : zie figuur op volgende slide - ‘de novo’ en ‘salvage pathway’ toepassing : bij co-transformatie met andere transgenen die men wou overbrengen : transfectie met 2 ongekoppelde DNA’s resulteerde in 90% co-transformanten ; m.a.w. TK+ cellen waren ook positief voor het niet-selecteerbare gen (uit analyse van deze regio bleek dat concatemeren tot 2 Mbp waren gevormd voor integratie optrad)
 Voorbeeld: Tk = thymidinekinasegen. historisch : experiment met muiscellen deficiënt in TK - te transformeren met. Herpes Simplex Virus (HSV) Tk gen => resultaat : activiteit herwonnen. transformanten te selecteren op HAT-medium (hypoxanthine - aminopterine - thymidine) : aminopterine blokkeert de de novo synthese van dTMP (en. dus van dTTP) voor achtergrond : zie figuur op volgende slide - ‘de novo’ en ‘salvage pathway’ toepassing : bij co-transformatie met andere transgenen die men wou overbrengen : transfectie met 2 ongekoppelde DNA’s resulteerde in 90% co-transformanten ; m.a.w. TK+ cellen waren ook positief voor het niet-selecteerbare gen (uit analyse van deze regio bleek dat concatemeren tot 2 Mbp waren gevormd voor integratie optrad)")
14
Endogene selectiemerkers : voorbeelden
DHFR Uridine

15
Dominante selectiemerkers (uitgebreide lijst in Primrose & Twyman, Tabel 12.3)
bruikbaar voor alle gevoelige cellijnen – nieuw fenotype meestal antibioticumresistentiemerkers van bacteriële oorsprong neomycinefosfotransferase – resistentie aan aminoglycosideantibiotica (kanamycine, neomycine, G418) – proteïnesynthese-inhibitoren selectiemerker wel onder transcriptieregulatie van eukaryotische elementen te plaatsen (bvb. HSV Tk promoter, SV40 ‘vroege’ promoter) ‘Kwantitatieve’ selectiemerkers (uitgebreide lijst in Primrose & Twyman, Tabel 12.4) stapsgewijze (random) amplificatie van de merker waardoor de cel ongevoelig wordt voor lage dosissen van de inhibitor/’drug’ toelaatbare concentratie – indicatie voor ‘kopijaantal’ transgen vb. dhfr-locus - dihydrofolaatreductase - methotrexaat is competitieve inhibitor voor deze enzymatische activiteit : schakelt de novo synthese van dUMP en dIMP uit toepassing: co-amplificatie van transgenen die gelijktijdig met DHFR geïntroduceerd werden - expressieniveau tot zeer hoog op te drijven Vb. methotrexaat-selectie voor grootschalige expressie : gradueel opdrijven van de concentratie methotrexaat ; kan tot maal de basis toxische concentratie tolereerbaar maken (o.a. toegepast voor productie van weefselplasminogeenactivator).
 – proteïnesynthese-inhibitoren. selectiemerker wel onder transcriptieregulatie van eukaryotische elementen. te plaatsen (bvb. HSV Tk promoter, SV40 ‘vroege’ promoter) ‘Kwantitatieve’ selectiemerkers (uitgebreide lijst in Primrose & Twyman, Tabel 12.4) stapsgewijze (random) amplificatie van de merker waardoor de cel ongevoelig wordt voor lage dosissen van de inhibitor/’drug’ toelaatbare concentratie – indicatie voor ‘kopijaantal’ transgen. vb. dhfr-locus - dihydrofolaatreductase - methotrexaat is competitieve inhibitor voor. deze enzymatische activiteit : schakelt de novo synthese van dUMP en dIMP uit. toepassing: co-amplificatie van transgenen die gelijktijdig met DHFR geïntroduceerd. werden - expressieniveau tot zeer hoog op te drijven. Vb. methotrexaat-selectie voor grootschalige expressie : gradueel opdrijven van de concentratie methotrexaat ; kan tot maal de basis toxische concentratie tolereerbaar maken (o.a. toegepast voor productie van weefselplasminogeenactivator).")
16
Repliconvectoren (viraal) - plasmidevectoren
Replicons Repliconvectoren (viraal) - plasmidevectoren Simian virus 40 (SV40) : eerste dierlijke virus dat onderzocht werd als mogelijke vector sequentie van het DNA genoom (5224 bp) bekend in 1978 Geïsoleerd uit de Rhesus aap : in 1960 uit culturen van niercellen, die werden gebruikt voor de aanmaak van het poliovaccin : controversieel toen bleek dat miljoenen mensen het hadden gekregen via het gecontamineerde vaccin. Papovaviridae (Papilloma, polyoma, vacuolating agent) (SV40 is een polyoma virus) SV40 is “slapend” en asymptomatisch in Rhesus apen. Bij de mens kan het een nierziekte veroorzaken. In andere species, vooral hamsters, veroorzaakt het tumoren. (Ook in de rat werd een tumormodel ontwikkeld.) Een formeel bewijs van een rol van SV40 bij tumor- vorming bij de mens ontbreekt (hoewel SV40-sequenties in heel wat tumoren gevonden werden). Groei op niercellen van de aap (AGMK, CV1, …) Structuur : 5243 bp (2 zeer kleine restrictiefragmentjes werden pas later ontdekt), circulair DNA chromatinestructuur (nucleosomen) ; rol van histon H1 (zie foto volgende slide) viruspartikel : icosahedraal, VP1, VP2, VP3 Genetische opbouw : "early" & "late" gebied, OR (replicatiefunctie) Lytische ontwikkeling versus cellulaire transformatie *
 - plasmidevectoren. Simian virus 40 (SV40) : eerste dierlijke virus dat onderzocht werd als mogelijke vector. sequentie van het DNA genoom (5224 bp) bekend in Geïsoleerd uit de Rhesus aap : in 1960 uit culturen van niercellen, die werden gebruikt voor de aanmaak van het poliovaccin : controversieel toen bleek dat miljoenen mensen het hadden. gekregen via het gecontamineerde vaccin. Papovaviridae (Papilloma, polyoma, vacuolating agent) (SV40 is een polyoma virus) SV40 is slapend en asymptomatisch in Rhesus apen. Bij de mens kan het een nierziekte. veroorzaken. In andere species, vooral hamsters, veroorzaakt het tumoren. (Ook in de rat. werd een tumormodel ontwikkeld.) Een formeel bewijs van een rol van SV40 bij tumor- vorming bij de mens ontbreekt (hoewel SV40-sequenties in heel wat tumoren gevonden werden). Groei op niercellen van de aap (AGMK, CV1, …) Structuur : 5243 bp (2 zeer kleine restrictiefragmentjes werden pas later ontdekt), circulair DNA. chromatinestructuur (nucleosomen) ; rol van histon H1 (zie foto volgende slide) viruspartikel : icosahedraal, VP1, VP2, VP3. Genetische opbouw : early & late gebied, OR (replicatiefunctie) Lytische ontwikkeling versus cellulaire transformatie. *")
17
* Vectoren voor dierlijke cellen, gebaseerd op SV40

18
* SV40 ori-gebied (details op volgende slide)
")
19
De origin voor SV40 DNA replicatie wordt voorgesteld door de ellips met erin een ruit
(tussen posities 5207 en 31). De belangrijkste elementen zijn een 27 bp palindroom en de aansluitende 17 AT-bp sequentie ("AT-block"). Als nulpunt (G) is de centrale positie genomen van het 27 bp palindroom (positie 1 ligt dus bij het volgende bp.) Het ligt aan de rechterzijde van een unieke BglI herkenningsplaats. cagaggccgaggcGgcctcggcctctg BglI GCCnnnnnGGC nummering Transcriptie van zowel het vroeg als laat gebied wordt geregeld door een (schijnbaar) bidirectionele promoter die de AT-block, 21 bp herhalingen, en de sequentie van de identische 72 bp gebieden omvat. (“schijnbaar” omdat deletie van sommige repeats een verschillend effect heeft op vroege versus late transcriptie.) Het kleinste ‘vroeg mRNA’ codeert voor groot-T antigen. Het grootste ‘vroeg mRNA’ voor klein-T antigen, hoewel het gehele vertaalde gebied (dus ook het stoptriplet) voor de positie van het intron ligt. Er is een (gemeenschappelijk) polyadenylatiesignaal, maar er is geen specifieke afstand vereist (m.a.w. bij deletiemutanten verschuift de positie naar links of rechts). Een groot intron zorgt voor het VP1 ‘laat mRNA’. De andere VP-eiwitten worden op grotere mRNAs gecodeerd (kleinere introns, diverse groottes).
. De belangrijkste elementen zijn een 27 bp palindroom en de. aansluitende 17 AT-bp sequentie ( AT-block ). Als nulpunt (G) is de centrale positie genomen van het 27 bp palindroom (positie 1 ligt dus. bij het volgende bp.) Het ligt aan de rechterzijde van een unieke BglI herkenningsplaats. cagaggccgaggcGgcctcggcctctg. BglI GCCnnnnnGGC nummering. Transcriptie van zowel het vroeg als laat gebied wordt geregeld door een (schijnbaar) bidirectionele promoter die de AT-block, 21 bp herhalingen, en de sequentie van de. identische 72 bp gebieden omvat. ( schijnbaar omdat deletie van sommige repeats een. verschillend effect heeft op vroege versus late transcriptie.) Het kleinste ‘vroeg mRNA’ codeert voor groot-T antigen. Het grootste ‘vroeg mRNA’ voor. klein-T antigen, hoewel het gehele vertaalde gebied (dus ook het stoptriplet) voor de positie. van het intron ligt. Er is een (gemeenschappelijk) polyadenylatiesignaal, maar er is geen. specifieke afstand vereist (m.a.w. bij deletiemutanten verschuift de positie naar links of rechts). Een groot intron zorgt voor het VP1 ‘laat mRNA’. De andere VP-eiwitten worden op grotere mRNAs gecodeerd (kleinere introns, diverse groottes).")
20
* N Transcriptiekaart van SV40. ORI ligt op positie 5243/0
Vroege transcriptie in tegenwijzerzin. 2 mRNAs door verschillende splicing (large-T + small-t) Late transcriptie in wijzerzin. 2 mRNAs, één voor VP1, andere voor VP2 + VP3 Gestippelde zones zijn de introns. N
 Late transcriptie in wijzerzin. 2 mRNAs, één voor VP1, andere voor VP2 + VP3. Gestippelde zones zijn de introns. N.")
21
‘Runaway’ polyoma replicons – replicerend, maar celdood
SV40 DNA 2 transcriptie-eenheden : vroeg en laat meerdere producten door alternatieve splicing Vroeg : regulatorische eiwitten Laat : capside-eiwitten Regulatie: regulatorische sequenties tussen vroege-late regio In regulatorische regio : vroege + late promoter, enhancer, ori Vroege eiwitten : t en T tumor antigenen T-antigen is vereist voor replicatie en de oncoproteïne-eigenschappen (Bemerk, en hou rekening met, de omgekeerde orientatie van de cirkel t.o.v. vorige figuur.) *
 *")
22
N Verloop van een SV40 infectie in permissieve en
in niet-permissieve cellen. Tot ongeveer 14 uur na infectie (vroege fase) is de de ontwikkeling gelijklopend. - In permissieve cellen volgt dan DNA replicatie, late eiwitsynthese, virusvorming en celdood. - In niet-permissieve cellen integreert het virale DNA in het cellulair DNA en kan “celtransformatie” optreden als expressie van groot-T antigen aanhoudt.
 is de. de ontwikkeling gelijklopend. - In permissieve cellen volgt dan DNA replicatie, late eiwitsynthese, virusvorming en celdood. - In niet-permissieve cellen integreert het virale. DNA in het cellulair DNA en kan celtransformatie optreden als expressie van groot-T antigen aanhoudt.")
23
N Infectieproces ontmanteling : 8 u
vroege fase: vroege mRNA's, vroege eiwitten : 4 u start replicatie + late fase : late mRNA's en eiwitten : vanaf +/- 24 u maturatie : inpakking en cellyse Aanmaak COS cellen : SV40 DNA gelineariseerd met BglI (3 bp uitstekende uiteinden) afsplitsing van de uitstekende uiteinden en ligeren => enkele bp in ORI verloren = ORI is inactief transformatie cellen : detectie T-antigen (op basis van fluorescentie) COS cellen : constitutieve expressie van T-antigen door het celgenoom => T-antigen wordt in trans voorzien. N niet N Naam COS : CV-1 van oorsprong (Origin), met SV40 CV-1 : een cellijn afgeleid van niercellen van de African green monkey aap. Immortalisatie door transformatie met het replicatie-deficiente SV40 DNA (COS cellen). Brengt wel nog het T-antigen tot expressie. Twee vormen van COS cellijnen : COS-1 (nog permissief voor lytische groei van SV40) en COS-7 (bevat defectieve genomen maar zet geen infectieve viruspartikels vrij). niet N
 afsplitsing van de uitstekende uiteinden en ligeren. => enkele bp in ORI verloren = ORI is inactief. transformatie cellen : detectie T-antigen (op basis van fluorescentie) COS cellen : constitutieve expressie van T-antigen door het celgenoom. => T-antigen wordt in trans voorzien. N. niet N. Naam COS : CV-1 van oorsprong (Origin), met SV40. CV-1 : een cellijn afgeleid van niercellen van de African green monkey aap. Immortalisatie door transformatie met het replicatie-deficiente SV40 DNA (COS cellen). Brengt wel nog het T-antigen tot expressie. Twee vormen van COS cellijnen : COS-1 (nog permissief voor lytische groei van SV40) en COS-7 (bevat defectieve genomen maar zet geen infectieve viruspartikels vrij). niet N.")
24
* Basisgegevens : Belangrijkste karakteristieken
deels N Belangrijkste karakteristieken * Basisgegevens : replicatie : ORI : minimaal 70 bp bi-directionele replicatie bij elke replicatieronde is opnieuw T-antigen nodig ORI : sluit promotoractiviteit in op beide strengen (tegengestelde richtingen) - vroege mRNA’s : 19S (T en t) - late mRNA’s : 16S (VP1) en 19S (VP2 en VP3) Beperkte inpakmogelijkheid : tot 110% van het genoom (30% deleties mogelijk) => dit limiteert vectorconstructie Bij pendelvectoren ( zie verder) moet rekening gehouden worden met zogenaamde “toxische” sequenties van pBR322 : dit zijn sequenties die de efficientie van transformatie van dierlijke cellen verminderen. Zij zijn gesitueerd in het gebied rond de nic/bom plaats van het plasmide.
 - vroege mRNA’s : 19S (T en t) - late mRNA’s : 16S (VP1) en 19S (VP2 en VP3) Beperkte inpakmogelijkheid : tot 110% van het genoom. (30% deleties mogelijk) => dit limiteert vectorconstructie. Bij pendelvectoren ( zie verder) moet rekening gehouden worden met zogenaamde. toxische sequenties van pBR322 : dit zijn sequenties die de efficientie van transformatie van dierlijke cellen verminderen. Zij zijn gesitueerd in het gebied rond de nic/bom plaats van het plasmide.")
25
Types SV40 vectoren : hoe, en welke, vectoren ontwikkeld op basis van SV40 elementen?
- transducerende vectoren (met helper virus) : inpakbaar, replicerend late replacement vectors : 2 soorten virions early replacement vectors : 2 soorten virions in COS cellen : zuivere virions - plasmide vectoren : niet-inpakbaar, replicerend gevolg van de vaststelling dat in COS cellen het viraal DNA repliceert, ook als het late gebied verwijderd is. Er worden dan uiteraard geen virions meer gevormd, dus geen plaques : er is bijgevolg een selectiemerker nodig. Er bleek ook geen grootte-beperking qua inserties te zijn. - passieve vectoren : niet-inpakbaar, niet-replicerend het DNA kan niet repliceren (niet-permissieve cellen), maar is alleen behoudbaar door integratie in het celgenoom. Dit werd mogelijk zodra selectiemerkers voor dierlijke cellen beschikbaar waren (en is dus bruikbaar in een groot aantal verschillende celtypes, ongeacht hun permissiviteit voor SV40). - pendelvectoren met E.coli plasmide : "toxisch" gebied (uit pBR322) te vermijden een SV40 replicon samengevoegd met een E. coli replicon. ook twee selectiemerkers vereist, één voor elk van beide waardcellen. N
 : inpakbaar, replicerend. late replacement vectors : 2 soorten virions. early replacement vectors : 2 soorten virions. in COS cellen : zuivere virions. - plasmide vectoren : niet-inpakbaar, replicerend. gevolg van de vaststelling dat in COS cellen het viraal DNA repliceert, ook als. het late gebied verwijderd is. Er worden dan uiteraard geen virions meer. gevormd, dus geen plaques : er is bijgevolg een selectiemerker nodig. Er bleek ook geen grootte-beperking qua inserties te zijn. - passieve vectoren : niet-inpakbaar, niet-replicerend. het DNA kan niet repliceren (niet-permissieve cellen), maar is alleen behoudbaar. door integratie in het celgenoom. Dit werd mogelijk zodra selectiemerkers voor dierlijke cellen beschikbaar waren (en is dus bruikbaar in een groot aantal verschillende celtypes, ongeacht hun permissiviteit voor SV40). - pendelvectoren met E.coli plasmide : toxisch gebied (uit pBR322) te vermijden. een SV40 replicon samengevoegd met een E. coli replicon. ook twee selectiemerkers vereist, één voor elk van beide waardcellen. N.")
26
Belangrijkste karakteristieken
Werken met : N (complementatie)
")
27
N Complementatie van SV40 transducerende vectoren met
defectieve helpers (ts-mutanten). Hetzij het "laat-DNA" (bovenaan) hetzij het "vroeg-DNA" (onderaan) is vervangen. Een temperatuur-gevoelige tsA mutatie geeft een mutant T, een tsB mutatie een mutant VP1. N nb. in deze en volgende figuur : vroegere nummering van het SV40 DNA vanaf de EcoRI knipplaats (in het VP1 gen) met het late gebied in wijzerzin.
. Hetzij het laat-DNA (bovenaan) hetzij het vroeg-DNA (onderaan) is vervangen. Een temperatuur-gevoelige. tsA mutatie geeft een mutant T, een tsB mutatie een mutant VP1. N. nb. in deze en volgende figuur : vroegere nummering van het. SV40 DNA vanaf de EcoRI. knipplaats (in het VP1 gen) met. het late gebied in wijzerzin.")
28
Complementatie door COS cellen, van een SV40 tranducerende
vector zonder groot-T gen. Het "laat-DNA" is vervangen door nieuwe insertsequenties. Na transfectie zijn alle eiwitten voor replicatie en verpakking aanwezig : T-antigen vanuit de COS cellen, VP1, 2 en 3 vanop de vector. niet N

29
Belangrijkste karakteristieken
Werken met : niet N N

30
N SV40 plasmidevectoren. Meestal pendelvectoren : bevatten ongeveer
2,3 kb pBR322 sequenties met de ori en het bla gen, voor klonering in E. coli. Het SV40 segment in de vector is het gebied van 5171 tot 270 (in wijzerzin) waarin de ORI ligt en ook de promoter-activiteit voor vroege transcriptie. Hierachter is een selectiemerker voorzien, waarachter de SV40 sequenties voor polyadenylering (rond positie 2500) liggen, zodat een actief mRNA gevormd wordt. pSV2 repliceert in COS cellen, maar niet in gewone apencellen waarin geen groot-T antigen aanwezig is. pSV3 repliceert wel in gewone apencellen, aangezien het groot-T groot-T gen ingebouwd is in de BamHI plaats van pSV2. pSV2 en pSV3 kunnen ook insereren in het celgenoom en permanente transformatie veroorzaken. Aldus groeit 1 cel op tot een kolonie in aanwezigheid van mycofenolzuur (een immunosuppressief agens). -gpt
 waarin de. ORI ligt en ook de promoter-activiteit voor vroege transcriptie. Hierachter is een selectiemerker voorzien, waarachter de SV40 sequenties voor polyadenylering (rond positie 2500) liggen, zodat een actief mRNA gevormd wordt. pSV2 repliceert in COS cellen, maar niet in. gewone apencellen waarin geen groot-T antigen aanwezig is. pSV3 repliceert wel in. gewone apencellen, aangezien het groot-T. groot-T gen ingebouwd is in de BamHI. plaats van pSV2. pSV2 en pSV3 kunnen ook insereren in het. celgenoom en permanente transformatie. veroorzaken. Aldus groeit 1 cel op tot een kolonie in aanwezigheid van. mycofenolzuur (een immunosuppressief agens). -gpt.")
31
Plasmidevectoren neo N Plasmidevector met expressie-eenheid op basis van het oncovirus RSV (Rous sarcoma virus) (RNA genoom) met LTR die specifieke integratie in het gastgenoom mogelijk maakt, evenals hoge genexpressie.
 (RNA genoom) met LTR die specifieke integratie in het gastgenoom mogelijk maakt, evenals hoge genexpressie.")
32
Niet-replicerende plasmiden
Samenvattend : deels * SV40 – virale vectoren eerste vectoren, virale vectoren gentransfer via transductie ; lytische infectie virions verplaatsing van vroege of late regio ; voorzie de ontbrekende regio in trans door co-introductie van een helpervirus vereenvoudiging door COS-cellijn : gen van T antigen genomisch in trans voorzien bij vectoren met uitgewisselde vroege regio nadeel : capaciteit viraal capside : max. ~ 2.5 kb te incorporeren ; SV40 ori op plasmiden voordeel : geen grootterestrictie enkel ori nodig replicatie in COS cellijn ; ori + gen T-antigen in permissieve cellen transiënt hoge expressie - aanmaak van grote hoeveelheden recombinant eiwit : bvb. in pcDNA3.1 ‘runaway’-karakter (groot aantal kopijen => grote hoeveelheden DNA) veroorzaakt celdood !! ontwikkeling van stabiele episomale vectoren mogelijk door regulatie van T antigen expressie (met conditionele promoter - temperatuursensitieve mutanten - e.d.) Niet-replicerende plasmiden niet-replicerende plasmiden blijven korte tijd extrachromosomaal in de cel aanwezig (lineair DNA wordt te snel afgebroken in zoogdiercellen, dus bij voorkeur cccDNA of covalent gesloten ocDNA). goed bruikbaar om snel met transiënte transformatie constructen uit te testen (regulatorische gebieden aangekoppeld aan een reportergen, aanmaak van een beperkte hoeveelheid van een nieuw recombinant eiwit (ter test, vooraleer een nieuwe stabiele cellijn te maken voor grootschalige productie). integratie in niet-permissieve cellen onder selectiedruk vanop het plasmide (geen co-transformatie nodig). Diverse vectoren zijn ontwikkeld (inclusief van SV40) vooral voor expressiedoeleinden. deels N
 veroorzaakt celdood. !! ontwikkeling van stabiele episomale vectoren mogelijk door regulatie van T antigen expressie (met conditionele promoter - temperatuursensitieve mutanten - e.d.) Niet-replicerende plasmiden. niet-replicerende plasmiden blijven korte tijd extrachromosomaal in de cel aanwezig (lineair. DNA wordt te snel afgebroken in zoogdiercellen, dus bij voorkeur cccDNA of covalent gesloten ocDNA). goed bruikbaar om snel met transiënte transformatie constructen uit te testen (regulatorische. gebieden aangekoppeld aan een reportergen, aanmaak van een beperkte hoeveelheid van een. nieuw recombinant eiwit (ter test, vooraleer een nieuwe stabiele cellijn te maken voor grootschalige. productie). integratie in niet-permissieve cellen onder selectiedruk vanop het plasmide (geen co-transformatie nodig). Diverse vectoren zijn ontwikkeld (inclusief van SV40) vooral voor expressiedoeleinden. deels N.")
33
Aantal voorbeelden enkele slides verder
* Transcriptie/translatie-vereisten Aantal voorbeelden enkele slides verder uitgebreidere beschrijving in Primrose & Twyman, Tabel 12.2) maximalisatie transgenexpressie gebruik van een sterke promoter Vb. adenovirale of vaccinia virus promoters SV40 vroege promoter + enhancer CMV (cytomegalovirus) vroege promoter !!! Activiteit is afhankelijk van celtype aanwezigheid van een intronsequentie : bevorderlijk voor expressie polyadenylatiesignaal (terminatoren) - voor correct 3’ uiteinde van het mRNA - toevoeging van > 100 adenylaat-residu’s - vaak poly(A) van SV40 vroege transcriptie-eenheid of muis b-globine mRNA - verhoogde stabiliteit 3’UTR en 5’ UTR regio’s van mRNA worden best (grotendeels) weggelaten in cDNA optimalisatie van transgen voor expressie : - consensus rond translatie-initiatieplaats - codonkeuze : potentieel tekort aan preferentiële codons : codonoptimalisatie voor de betrokken gastheer signaalsequentie voor secretorische pathways, bvb. Ig lichte keten N
 maximalisatie transgenexpressie. gebruik van een sterke promoter. Vb. adenovirale of vaccinia virus promoters. SV40 vroege promoter + enhancer. CMV (cytomegalovirus) vroege promoter. !!! Activiteit is afhankelijk van celtype. aanwezigheid van een intronsequentie : bevorderlijk voor expressie. polyadenylatiesignaal (terminatoren) - voor correct 3’ uiteinde van het mRNA. - toevoeging van > 100 adenylaat-residu’s. - vaak poly(A) van SV40 vroege transcriptie-eenheid of muis b-globine mRNA. - verhoogde stabiliteit. 3’UTR en 5’ UTR regio’s van mRNA worden best (grotendeels) weggelaten in cDNA. optimalisatie van transgen voor expressie : - consensus rond translatie-initiatieplaats. - codonkeuze : potentieel tekort aan preferentiële codons : codonoptimalisatie voor de. betrokken gastheer. signaalsequentie voor secretorische pathways, bvb. Ig lichte keten. N.")
34
Voorbeeld van expressieconstruct in een ‘runaway’ polyoma replicon
* Voorbeeld van expressieconstruct in een ‘runaway’ polyoma replicon N Zie ook hoofdstuk Expressievectoren en eiwitproductie voor nadere beschrijving van de diverse elementen BGH : “bovine growth hormone” sequentie pA : polyadenyleringssignaal Hoog expressieniveau in COS cellen (ORI tot 105 kopijen/cel). celdood ; beperkt aantal cellen zijn getranfecteerd neo als selectiemerker, tussen SV40-promoter en poly(A)-signaal PCMV = humane CMV (cytomegalovirus) promoter EK = enterokinase knipplaats voor verwijdering tags
. celdood ; beperkt aantal cellen. zijn getranfecteerd. neo als selectiemerker, tussen. SV40-promoter en poly(A)-signaal. PCMV = humane CMV (cytomegalovirus) promoter. EK = enterokinase knipplaats voor verwijdering tags.")
35
Ter illustratie van verdere ontwikkelingen met andere virussen
Algemene basisaspecten en overwegingen voor en bij gebruik van (andere) virussen voor gentransfer en expressie. Papillomavirussen (mens, rund, …) (wratten, …) Adenovirussen Herpesvirussen (EBV, e.a.) Baculovirus (bij insecten) Retrovirussen e.a. Verder ook : C. elegans (Caenorhabditis elegans - nematode) als modelorganisme) Zeer veel virussen en hun genomen werden onderzocht als mogelijkheid tot vectorontwikkeling en klonering, in functie van hun typische karakteristieken, gastcelspectrum, specifieke toepassingen, enz…
 virussen voor gentransfer en expressie. Papillomavirussen (mens, rund, …) (wratten, …) Adenovirussen. Herpesvirussen (EBV, e.a.) Baculovirus (bij insecten) Retrovirussen. e.a. Verder ook : C. elegans (Caenorhabditis elegans - nematode) als modelorganisme) Zeer veel virussen en hun genomen werden onderzocht als mogelijkheid tot vectorontwikkeling en klonering, in functie van hun typische karakteristieken, gastcelspectrum, specifieke toepassingen, enz…")
36
Virussen als gentransfervectoren - transducerende vectoren
Transductie : horizontale DNA-transfer via mechanisme van virusinfectie Gentransfer in vitro (celculturen) en in vivo (gentherapie toepassingen) Insertie van een transgen of omwisseling van virale sequenties voor een transgen Insertie door ligatie of homologe recombinatie Helper-onafhankelijk : kan zich autonoom repliceren + propageren Helper-afhankelijk : virale elementen moeten in trans voorzien worden co-infectie met helpervirus ; helperplasmide voorzien of een ‘complementaire cellijn’ (ook ‘packaging’ cellijn) Vaak gewenst : afwezigheid van virale coderende sequenties enkel amplicons overhouden (cis-elementen) voordelen : niet-cytotoxisch en grotere inserten mogelijk Keuze virale vector : afhankelijk van de te transformeren gastheer type expressie (stabiel - transiënt) hoeveelheid te verpakken DNA adenovirus - retrovirus - beperkte verpakkingscapaciteit (icosaëdrale hoofdjes) baculovirus - staafvormig - minder strikt
 en in vivo (gentherapie toepassingen) Insertie van een transgen of omwisseling van virale sequenties voor een transgen. Insertie door ligatie of homologe recombinatie. Helper-onafhankelijk : kan zich autonoom repliceren + propageren. Helper-afhankelijk : virale elementen moeten in trans voorzien worden. co-infectie met helpervirus ; helperplasmide voorzien of een ‘complementaire cellijn’ (ook ‘packaging’ cellijn) Vaak gewenst : afwezigheid van virale coderende sequenties enkel amplicons. overhouden (cis-elementen) voordelen : niet-cytotoxisch en grotere inserten mogelijk. Keuze virale vector : afhankelijk van de te transformeren gastheer. type expressie (stabiel - transiënt) hoeveelheid te verpakken DNA. adenovirus - retrovirus - beperkte verpakkingscapaciteit. (icosaëdrale hoofdjes) baculovirus - staafvormig - minder strikt.")
37
Expressie :

38
BPV – Bovine papillomavirus
Papilloma virussen : episomaal - structureel onstabiel BPV – Bovine papillomavirus Historisch - de eerste episomale vectoren (verre verwantschap met polyomavirussen) Kan muiscellen infecteren, zonder virus te produceren Kopijenaantal ~100 (geen integratie) Ook T antigen (replicatie + oncogene transformatie gastheercel) (vroegste deel van infectie similair aan SV40 infectie, latere meer complex) Vroege functies op 69% fragment - BPV69T selectiemerker neo - selectie op geneticine Vb. expressie humaan – b-globine Nadeel : recombinatie - deleties - zekere vorm van instabiliteit
 Kan muiscellen infecteren, zonder virus. te produceren. Kopijenaantal ~100 (geen integratie) Ook T antigen (replicatie + oncogene. transformatie gastheercel) (vroegste deel van infectie similair aan SV40 infectie, latere meer complex) Vroege functies op 69% fragment - BPV69T. selectiemerker neo - selectie op geneticine. Vb. expressie humaan – b-globine. Nadeel : recombinatie - deleties - zekere vorm van instabiliteit.")
39
BK virus (mens polyomavirus) JC (of JCV) is een ander voorbeeld ;
Papilloma virussen : human BK replicon ; episomaal - structureel onstabiel BK virus (mens polyomavirus) JC (of JCV) is een ander voorbeeld ; beide hebben ongeveer 70% homologie met SV40 DNA) Virus dat latente infectie veroorzaakt (infecteert meerdere celtypes) Behoud als episomaal replicon (geen verpakking in virions) Geen integratie => geen ‘positie-effect’ (zoals bij geïntegreerde transgenen) Relatief laag kopijenaantal = replicon interfereert niet met gastheercelgroei Plasmiden met ‘latente’ ori (geen inpakking in capsiden) Voordeel t.o.v. integratieve vectoren: geen integratiespecifieke beïnvloeding van expressie (m.a.w. geen ‘positie’-effect) Humaan BK polyomavirus –– kopijenaantal ~500 kan opgedreven orden tot ~ 9000 (door toenemende concentratie ‘antibiotica’ waartegen een resistentiegen aanwezig is, bvb. neoR) BK T-antigen kan voorzien worden in trans
 JC (of JCV) is een ander voorbeeld ; beide hebben ongeveer 70% homologie met SV40 DNA) Virus dat latente infectie veroorzaakt (infecteert meerdere celtypes) Behoud als episomaal replicon (geen verpakking in virions) Geen integratie => geen ‘positie-effect’ (zoals bij geïntegreerde transgenen) Relatief laag kopijenaantal = replicon interfereert niet met gastheercelgroei. Plasmiden met ‘latente’ ori (geen inpakking in capsiden) Voordeel t.o.v. integratieve vectoren: geen integratiespecifieke beïnvloeding. van expressie (m.a.w. geen ‘positie’-effect) Humaan BK polyomavirus –– kopijenaantal ~500. kan opgedreven orden tot ~ 9000 (door toenemende concentratie ‘antibiotica’ waartegen een resistentiegen aanwezig is, bvb. neoR) BK T-antigen kan voorzien worden in trans.")
40
Voorbeeld van een BKV vector (replicon)
BKV ori T antigen - functies

41
Epstein-Barr virus gebaseerde plasmiden – stabiele expressie
EBV : zeer stabiele replicatie in mens-cellen (+ mammalia in het algemeen) EBV - herpesvirus - dsDNA ~170 kb oorzaak van mononucleosis (‘klierkoorts’) episomaal replicon, ~ 1000 kopijen onder natuurlijke omstandigheden : alleen infectie van lymfocyten, maar na transfectie ook in tal van andere primaatcellen te behouden vereist voor episomale replicatie: latente oriP en trans-werkende regulator (EBNA1) EBV-gebaseerde expressievectoren – 2 tot 50 kopijen per cel selectie : ‘neomycine’ of ‘hygromycine’ selectiemerkers - stabiele aanwezigheid gebruik : episomale cDNA banken introductie van oriP in YACs in vitro circularisatie introductie in humane cellen
 EBV - herpesvirus - dsDNA ~170 kb. oorzaak van mononucleosis (‘klierkoorts’) episomaal replicon, ~ 1000 kopijen. onder natuurlijke omstandigheden : alleen infectie van lymfocyten, maar na. transfectie ook in tal van andere primaatcellen te behouden. vereist voor episomale replicatie: latente oriP en trans-werkende regulator (EBNA1) EBV-gebaseerde expressievectoren – 2 tot 50 kopijen per cel. selectie : ‘neomycine’ of ‘hygromycine’ selectiemerkers - stabiele aanwezigheid. gebruik : episomale cDNA banken. introductie van oriP in YACs. in vitro circularisatie introductie in humane cellen.")
42
Epstein-Barr virus gebaseerde plasmiden – stabiele expressie

43
Adenovirale vectoren voor ‘kortstondige’ transiënte expressie
Adenovirus genoom Adenovirus en adenovirus-afgeleide vectoren lineair, dsDNA genoom (~37 kb) 6 vroege transcriptie-eenheden (E) : virale replicatie 1 majeure late transcriptie-eenheid (MLT) : capside-eiwitten veel gebruikt als gentransfer- & genexpressievector & attractief voor faagtherapie (efficiënte opname – neg./inflammatoire respons – immuunsysteem) voordelen: stabiliteit, hoge capaciteit vreemd DNA, hoge titer (1011 pfu/ml), breed gastheerspectrum transiënte expressie in delende cellen – geen efficiënte integratie
 6 vroege transcriptie-eenheden (E) : virale replicatie. 1 majeure late transcriptie-eenheid (MLT) : capside-eiwitten. veel gebruikt als gentransfer- & genexpressievector & attractief voor. faagtherapie (efficiënte opname – neg./inflammatoire respons – immuunsysteem) voordelen: stabiliteit, hoge capaciteit vreemd DNA, hoge titer (1011 pfu/ml), breed gastheerspectrum. transiënte expressie in delende cellen – geen efficiënte integratie.")
44
Adenovirale vectoren:
- eerste generatie : replicatiedeficiënt : ‘E1 replacement vectors’ - kloneercapaciteit cfr. ‘uitgewisseld’ gedeelte ~7 kb - productie in de humane embryonale niercellijn 293 die de linker 11% van het adenovirus DNA bevat complementatie van functies in trans - ook andere ‘E’-regio’s verwijderd eventueel grotere inserties (~10 kb capaciteit) - nadelen : virale expressie – cytotoxiciteit eventueel recombinatie met geïntegreerde complementerende sequenties - enkel amplicon overhouden - grote capaciteit (~ 37 kb) - minimaal cytotoxisch langere expressie - nadeel : geen cellijn die alle functies in trans voorziet verpakking vereist helpervirus risico van contaminatie
 - nadelen : virale expressie – cytotoxiciteit. eventueel recombinatie met geïntegreerde. complementerende sequenties. - enkel amplicon overhouden. - grote capaciteit (~ 37 kb) - minimaal cytotoxisch langere expressie. - nadeel : geen cellijn die alle functies in trans. voorziet verpakking vereist helpervirus. risico van contaminatie.")
45
Vectorafgeleiden van het adeno-geassocieerd virus - integratief
Adeno-associated virus (AAV) AAV : ontdekt als contaminant in een adenoviraal preparaat ssDNA virus - Parvoviridae AAV-genoom : ~5 kb rep (replicatie) en cap (capside) genen geflankeerd door 145 nt omgekeerde herhalingen van nature replicatiedeficiënt - vereist ander virus (bvb. adenovirus, herpesvirus) voor lytische infectiecyclus zonder ‘helper’ : integratie in het genoom (constante locus - chromosoom 19) latent provirus ‘rescue’ door o.a. een adenovirus-infectie noodzaak van helperinfectie = controle over virusreplicatie – veiligheid bij gentherapietoepassing (samen met specifieke integratie)
 AAV : ontdekt als contaminant in. een adenoviraal preparaat. ssDNA virus - Parvoviridae. AAV-genoom : ~5 kb. rep (replicatie) en cap (capside) genen geflankeerd door 145 nt. omgekeerde herhalingen. van nature replicatiedeficiënt - vereist. ander virus (bvb. adenovirus, herpesvirus) voor lytische infectiecyclus. zonder ‘helper’ : integratie in het genoom (constante locus - chromosoom 19) latent provirus ‘rescue’ door o.a. een adenovirus-infectie. noodzaak van helperinfectie = controle over virusreplicatie – veiligheid bij. gentherapietoepassing (samen met specifieke integratie)")
46
Rep proteïnen interfereren met endogene promoters cytotoxisch effect
Vectorontwikkeling : Rep proteïnen interfereren met endogene promoters cytotoxisch effect deletie rep en cap genen LTR enige noodzakelijke elementen voor replicatie, transcriptie, provirale integratie (eventueel niet plaatsspecifiek in afwezigheid van Rep-eiwitten) en ‘rescue’. transfectie van AAV-afgeleid construct : LTR + transgen (in plasmidevector) + AAV-functies (helperplasmide) - transfectie-gebaseerd adenoviraal helperplasmide (virusvorming) affiniteitschromatografie voor opzuivering AAV-partikels – zoniet contaminatie met adenoviruspartikels in principe beperkte capaciteit verpakking vreemd DNA - aflevering van beperkte beperkte hoeveelheid DNA omzeilen via co-introductie van 2 segmenten op 2 vectoren vectorconcatemerisatie in doelwitcel
 en ‘rescue’. transfectie van AAV-afgeleid construct : LTR + transgen (in plasmidevector) + AAV-functies (helperplasmide) - transfectie-gebaseerd adenoviraal helperplasmide (virusvorming) affiniteitschromatografie voor opzuivering AAV-partikels – zoniet contaminatie. met adenoviruspartikels. in principe beperkte capaciteit verpakking vreemd DNA - aflevering van beperkte. beperkte hoeveelheid DNA omzeilen via co-introductie van 2 segmenten. op 2 vectoren vectorconcatemerisatie in doelwitcel.")
47
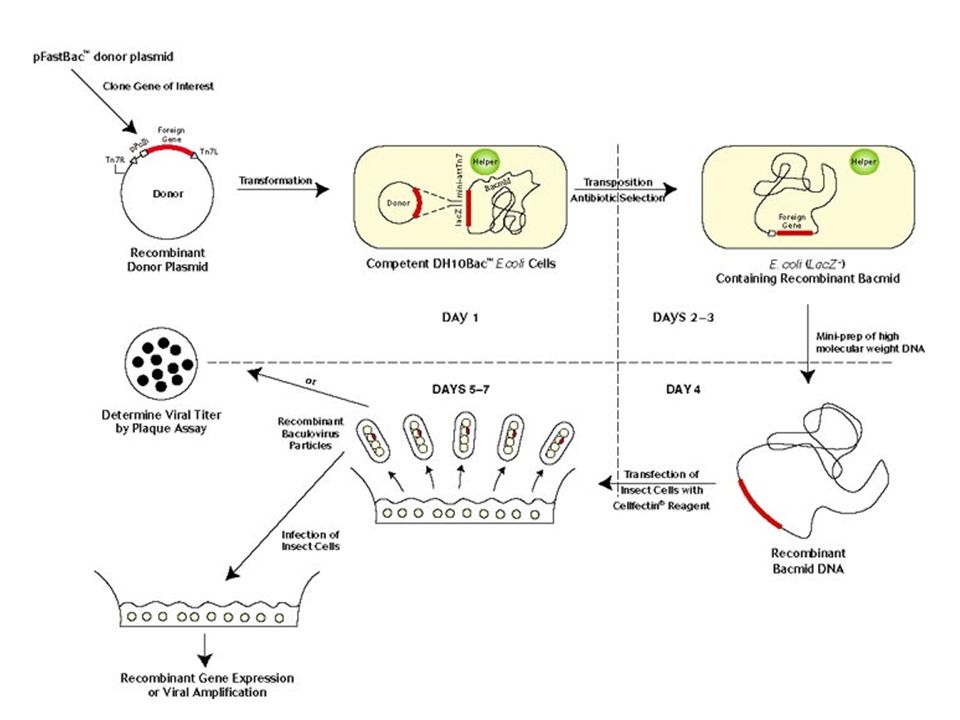
Baculovirus-afgeleide vectoren: hoog expressieniveau in insectcellijnen
staafvormig capside groot dsDNA genoom gastheer: insekten bepaalde baculovirussen : polyhedrosis virussen – productie van ‘nuclear occlusion bodies’ virions omgeven door eiwitlaag (polyhedrine) – extra bescherming polyhedrine productie - hoog - sterke promoter polyhedrine is niet essentieel (voor geslaagde infectie van cellijnen) polyhedrine-gen uit te wisselen voor een transgen = polyhedrine ‘replacement’ vectoren (hoog expressieniveau mg/106 cellen) Meest toegepast : ‘Autographa californica multiple nuclear polyhedrosis virus’ (AcMNPV) eiwitexpressie in cellijnen (bvb. Sf9, Sf21) ‘Bombyx mori nuclear polyhedrosis virus’ (BmNPV) infectie zijderups - recombinante eiwitexpressie in organisme
 – extra bescherming. polyhedrine productie - hoog - sterke promoter. polyhedrine is niet essentieel (voor geslaagde infectie van cellijnen) polyhedrine-gen uit te wisselen voor een transgen. = polyhedrine ‘replacement’ vectoren. (hoog expressieniveau - 1 mg/106 cellen) Meest toegepast : ‘Autographa californica multiple nuclear polyhedrosis virus’ (AcMNPV) eiwitexpressie in cellijnen (bvb. Sf9, Sf21) ‘Bombyx mori nuclear polyhedrosis virus’ (BmNPV) infectie zijderups - recombinante eiwitexpressie in organisme.")
48
Baculovirus Endocytosis van baculovirus ‘replacement vectors’ uitwisseling van polyhedrine-gen : screening van recombinanten : * Wild-type virus productie occlusion bodies opaque plaques => recombinanten geen occlusion bodies heldere plaques * E. coli lacZ gen in leesraam blauw-wit onderscheid * ‘Green fluorescent eiwit’ (GFP) in leesraam
 in leesraam.")
49
‘polyhedrine replacement vectors’
Opmerking bij deze illustratie meer succesvolle systemen werden ontwikkeld met een grotere deletie dan enkel het polyhedrine-gen

50
DNA en vector met transgen en homologe regio’s
Klonering door homologe recombinatie tussen polyhedrine-gedeleteerd viraal DNA en vector met transgen en homologe regio’s * nadeel bij beperkte deletie : lage frequentie recombinanten (0,5-5 %) * oplossing met gelineariseerd DNA door homologe recombinatie in een wild-type baculovirus met grote deleties (defectief); circulariteit enkel te herstellen door homologe recombinatie met een vector waarin het target transgen + replicon : wordt replicatie competent. leefbaar, recombinant genoom tot 90% recombinanten * bacmid : gebaseerd op elementen uit een fusie van een BAC vector met een baculovirus (gebaseerd op plaats-specifieke transpositie) (figuur volgende slide) Glycosylatie : - is mogelijk : echter : glycosylatiereactieweg verschillend tussen insecten en zoogdieren => mogelijk onjuiste glycosylatie immunogeen recombinant eiwit - oplossing : cellijnen die zoogdiereigen glycosylatie uitvoeren of co-expressie van geschikte glycosylatie-enzymen - vb. expressie van plasminogeenactivator (gegalactosyleerd) => de Sf9-cellijn brengt het 1,4-galactosyltransferase tot expressie (onder controle van een vroege baculoviruspromoter, dus induceerbaar door baculovirus infectie). Een baculovirus vector die een plasminogeenactivator transgen tot expressie brengt wordt hierdoor correct geglycosyleerd.
 * oplossing met gelineariseerd DNA door homologe recombinatie in. een wild-type baculovirus met grote deleties (defectief); circulariteit enkel. te herstellen door homologe recombinatie met een vector waarin het. target transgen + replicon : wordt replicatie competent. leefbaar, recombinant genoom tot 90% recombinanten. * bacmid : gebaseerd op elementen uit een fusie van een BAC vector met een baculovirus (gebaseerd op plaats-specifieke transpositie) (figuur volgende slide) Glycosylatie : - is mogelijk : echter : glycosylatiereactieweg verschillend tussen insecten. en zoogdieren => mogelijk onjuiste glycosylatie. immunogeen recombinant eiwit. - oplossing : cellijnen die zoogdiereigen glycosylatie uitvoeren. of co-expressie van geschikte glycosylatie-enzymen. - vb. expressie van plasminogeenactivator (gegalactosyleerd) => de Sf9-cellijn brengt het 1,4-galactosyltransferase tot expressie (onder controle van een vroege baculoviruspromoter, dus induceerbaar door baculovirus. infectie). Een baculovirus vector die een plasminogeenactivator. transgen tot expressie brengt wordt hierdoor correct geglycosyleerd.")
52
Herpesvirus vectoren – latent – mogelijk lange termijn transgenexpressie
Herpesvirussen - groot dsDNA - o.a. EBV, HSV, Varicella zoster HSV-1 gebaseerde transducerende vectoren na transfer door transfectie – compatibel met vele cellijnen evenwel : via transductie - voor gentherapie - neurotopisch = gericht naar zenuwcellen groot virus - transgenklonering via homologe recombinatie in getransfecteerde cellen alternatief : plasmide-gebaseerde ampliconvectoren - enkel cis-elementen (replicatie & verpakking) + verpakkingssystemen voorzien in trans gentransfer in vivo naar neuronen
 + verpakkingssystemen voorzien in trans. gentransfer in vivo naar neuronen.")
53
Retrovirale vectoren – efficiënte genomische integratie
Retrovirussen : RNA-virussen die repliceren via DNA-intermediair Integratie in genoom gastheercel Vectorontwikkeling - belangrijke kenmerken: - sommige acuut oncogeen - replicatie-deficiënt - geen gastheerafdoding - continue productie - vaak sterke promoters - hoge eiwitexpressie (mogelijk induceerbaar) - sommige breed celspectrum - geschikt voor vectorontwikkeling door beperkte genoomgrootte (in vitro-manipulaties eenvoudig) - hoge titers - efficiënte infectie Nadelen: velen – enkel efficiënte infectie van delende cellen beperkt qua gentherapie-toepassingen Lentiviridae (o.a. HIV) wel mogelijkheid van infectie van niet-delende cellen
 - sommige breed celspectrum. - geschikt voor vectorontwikkeling door beperkte genoomgrootte. (in vitro-manipulaties eenvoudig) - hoge titers - efficiënte infectie. Nadelen: velen – enkel efficiënte infectie van delende cellen. beperkt qua gentherapie-toepassingen. Lentiviridae (o.a. HIV) wel mogelijkheid van infectie van niet-delende cellen.")
54
Oncoretrovirussen - genoom
Boven: geïntegreerd provirus – LTR PB = primer binding sites - virale replicatie Onder : verpakt RNA Geïntegreerd genoom bevat gag, pol, env genen : gag structurele eiwitten pol reverse transcriptase env enveloppe-eiwitten Viraal RNA afgeschreven van promoter in linker LTR en stopt na polyadenyleringssignaal in rechter LTR. Ook capping. Gag-pol fusieproteïne processing tot meerdere polypeptiden Deel RNA splicing gag-pol en translatie env Twee kopijen RNA genoom verpakt + reverse transcriptase & integrase

Verwante presentaties
>")









.>")








