Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
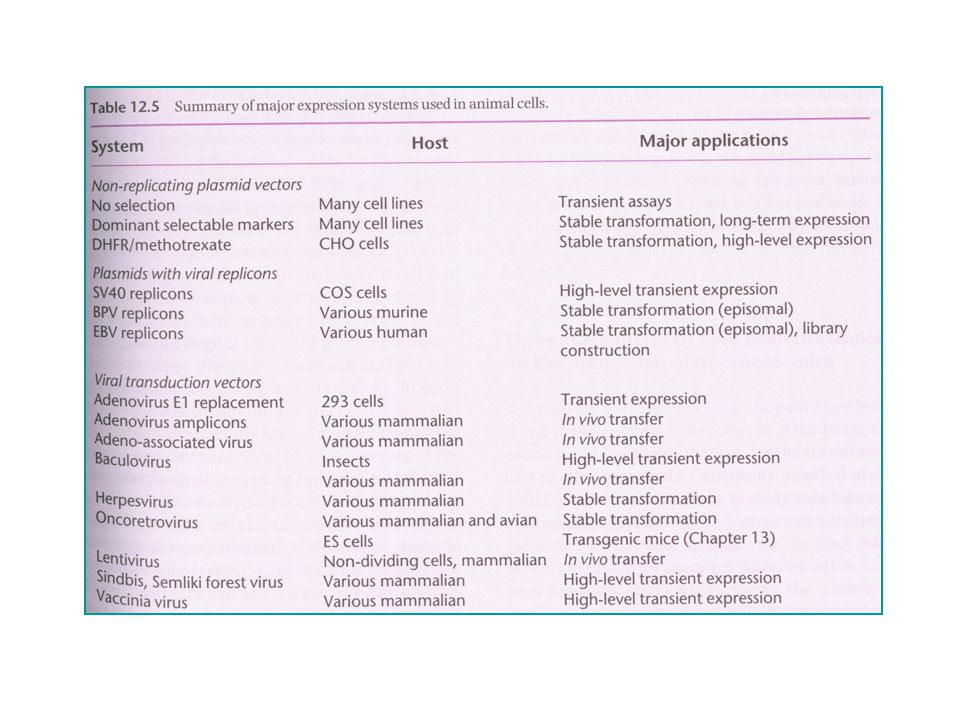
Gentransfer naar dierlijke cellen
(DNA opname : Primrose & Twyman, pp ) Belang: studie genfunctie – genregulatie aanmaak recombinante eiwitten – juiste posttranslationele modificaties manipuleren van endogeen genoom - gentherapie commercieel: synthese, o.a. vaccins, hormonen in zoogdier-celculturen gene-delivery voor dierlijke cellen in vivo versus in vitro : gentherapie
 Belang: studie genfunctie – genregulatie. aanmaak recombinante eiwitten – juiste posttranslationele modificaties. manipuleren van endogeen genoom - gentherapie. commercieel: synthese, o.a. vaccins, hormonen in zoogdier-celculturen. gene-delivery voor dierlijke cellen in vivo versus in vitro : gentherapie.")
2
Gentransfer naar dierlijke cellen
4 hoofdstrategiën – horizontale gentransfer Chemische - Fysische “transfectie”/transformatie Selectiemerkers Plasmidevectoren – dierlijke + bacteriële componenten DNA-”delivery” met bacteriële vectoren Virussen als gentransfer vectoren Hoofdstrategiën transformatie “niet-biologisch” (transfectie) Transformatie - fysisch Transformatie - chemisch - endocytose Transductie – virus-gemediëerd ‘Bactofection’ – bacterie-gemediëerd “biologisch” Transformatie
 Transformatie - fysisch. Transformatie - chemisch - endocytose. Transductie – virus-gemediëerd. ‘Bactofection’ – bacterie-gemediëerd. biologisch Transformatie.")
3
DNA transfer Chemische methoden – transformatie (transfectie) complex zodanig dat interactie met celmembraan bevorderd wordt vb. Positieve lading opname door endocytose Lipofiel complex membraanfusie DNA in cytoplasma Fysische methoden – transformatie (transfectie) door fysische kracht introductie van ‘naakt’ DNA vb. micro-injectie – “particle bombardment” – electroporatie – ultrasone behandeling
 door fysische kracht introductie van ‘naakt’ DNA. vb. micro-injectie – particle bombardment – electroporatie. – ultrasone behandeling.")
4
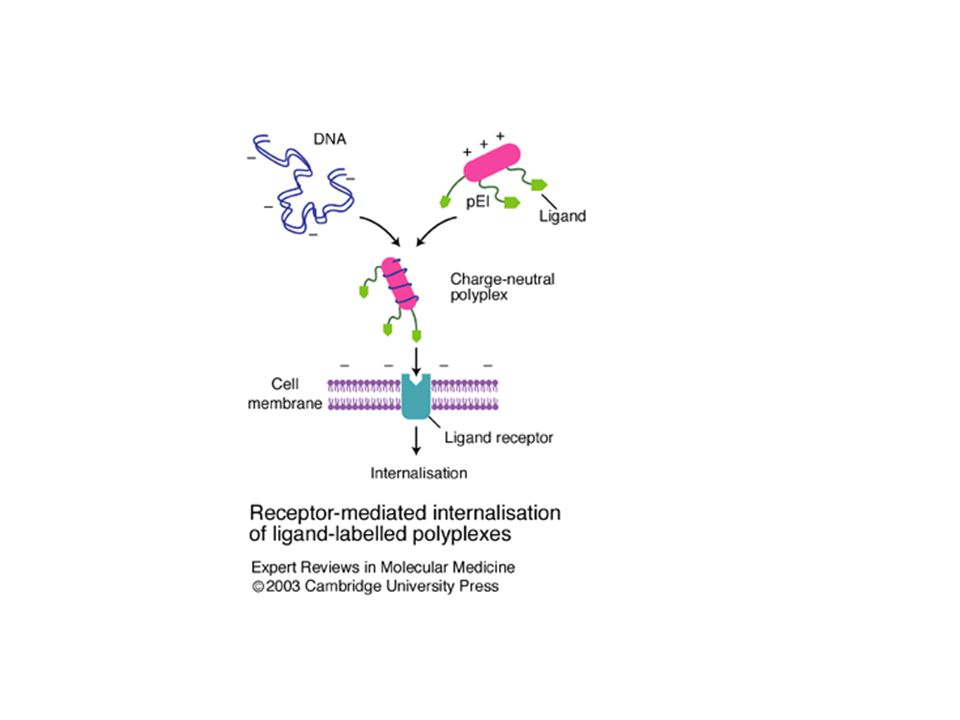
Chemische methoden – transformatie (transfectie)
Calciumfosfaat - DNA/calciumfosfaat co-precipitaat afzetting op de membraan en daarna internalisatie – endocytosis - naar nucleus voor expressie - “coating” van cellen nodig; cellen in monolayers of suspensies; niet geschikt voor cellen in “clumps” - 1-2% van de cellen nemen DNA op – slechts beperkte stabiele transformatie - optimalisatie via specifieke buffersystemen voor trage precipitatie en optimale balans tussen DNA, calciumfosfaat, buffer/pH Polyplexen (polykationisch polymeer in complex met DNA) - “probleemcellen” niet te transformeren via calciumfosfaat - polykationische componenten die spontaan oplosbare complexen vormen (“polyplexen”) – elektrostatische interactie met DNA - DEAE (diethylamino ethyldextraan) – polykationisch koolhydraat
 - probleemcellen niet te transformeren via calciumfosfaat. - polykationische componenten die spontaan oplosbare complexen. vormen ( polyplexen ) – elektrostatische interactie met DNA. - DEAE (diethylamino ethyldextraan) – polykationisch koolhydraat.")
5
Diethylaminoethyl dextran
DEAE Dextran

6
Chemische methoden – transformatie (transfectie)
Polyplexen - DEAE (diethylamino ethyldextraan) – polykationisch koolhydraat moeilijk voor stabiele transformatie - nieuwe generatie polykationische componenten -- Tabel 12.1 e.g., poly-L-lysine, spermine, synthetische polyamines – veel aminogroepen – positieve ladingen polyethylenimines, dendrimers - ook voor in vivo gentransfer: controle over genexpressie en gen’delivery’ via specifieke polymeren met veranderende eigenschappen vb. o.i.v. temperatuursregulatie
 – polykationisch koolhydraat. moeilijk voor stabiele transformatie. - nieuwe generatie polykationische componenten -- Tabel e.g., poly-L-lysine, spermine, synthetische polyamines – veel aminogroepen – positieve ladingen. polyethylenimines, dendrimers. - ook voor in vivo gentransfer: controle over genexpressie en gen’delivery’ via specifieke polymeren met veranderende eigenschappen vb. o.i.v. temperatuursregulatie.")
8
Ter illustratie

9
Ter illustratie

10
Chemische methoden – transformatie (transfectie)
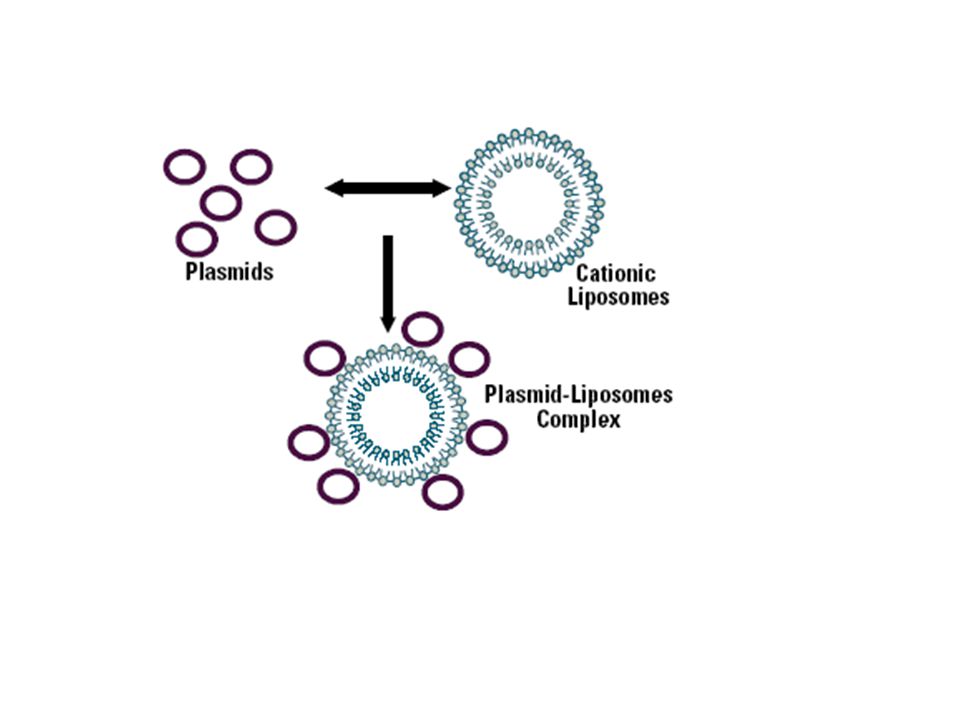
Liposomen en lipoplexen - verpakking in liposoom (‘fusogene’ fosfolipidevesikels) membraanfusie (eerste voorbeeld : Schaffner in 1980 : fusie van bacteriële protoplasten met dierlijke cellen om DNA binnen te brengen) - liposomen = artificiële fosfolipidevesikels - kationische/neutrale lipidemengsels vormen stabiele complexen met DNA = lipoplexen - mogelijkheid tot transformatie van grote DNA’s - “zachte” weg - hoge efficiëntie van transformatie (tot 90% van de cellen) – zowel transiënt als stabiel - transformatie m.b.v. liposomen = lipofectie - mogelijkheid transformatie van cellen in levende organismen na injectie in doelwitweefsels (gen’delivery’) – optimalisatie via insertie van virale eiwitten die celfusie promoten of targetting naar specifieke cellen
 membraanfusie. (eerste voorbeeld : Schaffner in 1980 : fusie van bacteriële protoplasten met dierlijke cellen om DNA. binnen te brengen) - liposomen = artificiële fosfolipidevesikels. - kationische/neutrale lipidemengsels vormen stabiele complexen met DNA. = lipoplexen. - mogelijkheid tot transformatie van grote DNA’s - zachte weg. - hoge efficiëntie van transformatie (tot 90% van de cellen) – zowel transiënt. als stabiel. - transformatie m.b.v. liposomen = lipofectie. - mogelijkheid transformatie van cellen in levende organismen na injectie in. doelwitweefsels (gen’delivery’) – optimalisatie via insertie van virale. eiwitten die celfusie promoten of targetting naar specifieke cellen.")
12
Liposoom Kationisch liposoom In interactie met DNA = lipoplex

13
Gentransfer naar dierlijke cellen
Fysische methoden – transformatie (transfectie) Elektroporatie - transiënte vorming van poriën (nm-grootte) - blootstelling elektrische puls – intensiteit/tijd – empirische optimalisatie - eenvoudig – reproduceerbaar - mogelijkheid in vivo gentransfer – oppervlakkig via elektroden of naaldelek- troden Ultrasone transfectie snel oscillerende probe (tip sonicator) ‘inklappen’ van membranen - ‘cavities’ – transiënte doorgang van DNA door membranen zowel in vitro als in vivo gen’delivery’
 Elektroporatie. - transiënte vorming van poriën (nm-grootte) - blootstelling elektrische puls – intensiteit/tijd – empirische optimalisatie. - eenvoudig – reproduceerbaar. - mogelijkheid in vivo gentransfer – oppervlakkig via elektroden of naaldelek- troden. Ultrasone transfectie. snel oscillerende probe (tip sonicator) ‘inklappen’ van membranen - ‘cavities’ – transiënte doorgang van DNA. door membranen. zowel in vitro als in vivo gen’delivery’")
14
Fysische methoden – transformatie (transfectie)
Micro-injectie directe transfer tot in de cel enkel toepasbaar voor een aantal celle meestal toegepast op grote celtypes, e.g. oocyten in vivo en in vitro “Particle bombardment” - directe transfer tot in de cel - initieel ontwikkeld voor transformatie van planten - kleine metalen (gouden) partikels worden gecoat met DNA en m.b.v. kracht geïntroduceerd
 partikels worden gecoat met DNA en m.b.v. kracht geïntroduceerd.")
15
Transformatie/transfectie – DNA in de cel
Transformatie in twee ‘stadia’: introductie van DNA in de cel (transfectie) incorporatie in de nucleus (veelal door integratie in het gastheerchromosoom) Transformatie: Kan tijdelijke toestand zijn = transiënte transformatie/transfectie – effect kortstondig Integratie in het genoom – overerfbaar – stabiele transgene cellijn Integratie = inefficiënt selectie vereist Merkers Drie types selectiemerkers in dierlijke cellen Endogene selectiemerkers – bruikbaar bij mutante cellijnen Dominante selectiemerkers ‘Kwantitatieve’ selectiemerkers
 incorporatie in de nucleus (veelal door integratie in het gastheerchromosoom) Transformatie: Kan tijdelijke toestand zijn = transiënte transformatie/transfectie – effect kortstondig. Integratie in het genoom – overerfbaar – stabiele transgene cellijn. Integratie = inefficiënt selectie vereist. Merkers. Drie types selectiemerkers in dierlijke cellen. Endogene selectiemerkers – bruikbaar bij mutante cellijnen. Dominante selectiemerkers. ‘Kwantitatieve’ selectiemerkers.")
16
Endogene selectiemerkers
Overzicht Tabel 12.2 Voorbeeld: Tk = thymidinekinasegen Historisch : experiment met muiscellen deficiënt in TK – te transformeren met Herpes Simplex Virus (HSV) Tk gen – resultaat : activiteit herwonnen transformanten te selecteren op HAT-medium (hypoxanthine – aminopterine – thymidine) : aminopterine blokkeert de de novo synthese van dTMP (en dus van dTTP) Achtergrond – figuur 12.3 (volgende slide) – ‘de novo’ en ‘salvage pathway’ Toepassing: bij co-transformatie met andere transgenen die men wou overbrengen Transfectie met 2 ongekoppelde DNA’s in 90% cotransformanten TK+ cellen ook positief voor niet-selecteerbaar gen (in zelfde regio : concatemeren tot 2 Mbp gevormd, vooraleer integratie)
 Tk gen – resultaat : activiteit herwonnen. transformanten te selecteren op HAT-medium (hypoxanthine – aminopterine – thymidine) : aminopterine blokkeert de de novo synthese van dTMP (en dus van dTTP) Achtergrond – figuur 12.3 (volgende slide) – ‘de novo’ en ‘salvage pathway’ Toepassing: bij co-transformatie met andere transgenen die men wou overbrengen. Transfectie met 2 ongekoppelde DNA’s in 90% cotransformanten. TK+ cellen ook positief voor niet-selecteerbaar gen. (in zelfde regio : concatemeren tot 2 Mbp gevormd, vooraleer integratie)")
17
Endogene selectiemerkers

18
Dominante selectiemerkers
Bruikbaar voor alle gevoelige cellijnen – nieuw fenotype Meestal antibioticumresistentiemerkers van bacteriële oorsprong Neomycinefosfotransferase – resistentie aan aminoglycosideantibiotica (kanamycine, neomycine, G418) – proteïnesynthese-inhibitoren Selectiemerker wel onder transcriptieregulatie van eukaryote elementen plaatsen (bvb. HSV Tk promoter, SV40 ‘vroege’ promoter) Overzicht: Tabel 12.3
 – proteïnesynthese-inhibitoren. Selectiemerker wel onder transcriptieregulatie van eukaryote elementen. plaatsen (bvb. HSV Tk promoter, SV40 ‘vroege’ promoter) Overzicht: Tabel")
19
‘Kwantitatieve’ selectiemerkers
stapsgewijze (random) amplificatie van de merker waardoor de cel ongevoelig wordt voor lage dosissen van de inhibitor/’drug’ toelaatbare concentratie – indicatie voor ‘kopijaantal’ transgen Vb. dhfr –locus - dihydrofolaatreductase – methotrexaat is competitieve inhibitor voor deze enzymatische activiteit Toepassing: co-amplificatie van transgenen die gelijktijdig met Dhfr geïntroduceerd werden – expressieniveau : hoog Vb. Methotrexaat-selectie voor grootschalige expressie van o.a. weefselplasminogeenactivator Overzicht: Tabel 12.4
 amplificatie van de merker waardoor de cel ongevoelig wordt voor lage dosissen van de inhibitor/’drug’ toelaatbare concentratie – indicatie voor ‘kopijaantal’ transgen. Vb. dhfr –locus - dihydrofolaatreductase – methotrexaat is competitieve. inhibitor voor deze enzymatische activiteit. Toepassing: co-amplificatie van transgenen die gelijktijdig met Dhfr geïntroduceerd werden – expressieniveau : hoog. Vb. Methotrexaat-selectie voor grootschalige expressie van o.a. weefselplasminogeenactivator. Overzicht: Tabel")
20
Transcriptie/translatie-vereisten
Box 12.2 maximalisatie transgenexpressie gebruik van een sterke promoter Vb. Adenovirale, vaccinia virus promoters SV40 vroege promoter + enhancer CMV vroege promoter ! Activiteit afhankelijk van celtype aanwezigheid van een intronsequentie: bevorderlijk voor expressie polyadenylatiesignaal (terminatoren) – voor correct 3’ uiteinde van mRNA Toevoeging van > 100 adenylaat-residu’s Vaak poly(A) van SV40 vroege transcriptie-eenheid of muis b-globine Verhoogde stabiliteit 3’UTR en 5’ UTR regio’s van mRNA worden best weggelaten in cDNA optimalisatie van transgen voor expressie: consensus rond translatie- initiatieplaats + codonkeuze – potentieel tekort aan preferentiële codons : codonoptimalisatie voor de betrokken gastheer signaalsequentie voor secretorische pathways, bvb. Ig lichte keten
 – voor correct 3’ uiteinde van mRNA. Toevoeging van > 100 adenylaat-residu’s. Vaak poly(A) van SV40 vroege transcriptie-eenheid of muis b-globine. Verhoogde stabiliteit. 3’UTR en 5’ UTR regio’s van mRNA worden best weggelaten in cDNA. optimalisatie van transgen voor expressie: consensus rond translatie- initiatieplaats + codonkeuze – potentieel tekort aan preferentiële. codons : codonoptimalisatie voor de betrokken gastheer. signaalsequentie voor secretorische pathways, bvb. Ig lichte keten.")
21
Plasmidevectoren – repliconvectoren - transfectie
Replicons Plasmidevectoren – repliconvectoren - transfectie Eerste experimenten : complexe mengsels van DNA of afzonderlijke genseq. Later : gebruik van plasmidevectoren : Voordelen : - gemak in vitro manipulaties - introductie van modulaire elementen (e.g. promotersequenties) die onafhankelijk van het transgen bruikbaar zijn - selectiemerker op het construct omzeilt de noodzaak van co-transformatie (transgen en selectiemerker gekoppeld) - mogelijkheid tot constructie pendelvectoren - vereisen geen integratie in het genoom - episomaal stabiel
 die. onafhankelijk van het transgen bruikbaar zijn. - selectiemerker op het construct omzeilt de noodzaak van. co-transformatie (transgen en selectiemerker gekoppeld) - mogelijkheid tot constructie pendelvectoren - vereisen geen integratie. in het genoom - episomaal stabiel.")
23
Plasmidevectoren neo

24
Niet-replicerende plasmiden
Toepassing in transiënte transformatie (zonder selectiemerker) zie ook Tabel voor eerste overzicht Transiënte expressie - transiënte evaluatie van genexpressie / eiwitexpressie testen van een grote reeks constructen in parallel - geen grootschalige productie. analyse van regulatorische elementen (Box 12.1) (reportergenen !) Hoe ? afwezigheid van functionele ori - zolang plasmide stabiel - expressie (’korte’ tijd = 1 of 2 dagen) ‘Runaway’ polyoma replicons – replicerend, maar celdood Transiënt - episomaal Ophoping van grote hoeveelheden DNA in korte tijd Vb. Simian virus SV40 en murine (muis) polyomavirus SV40, eerste dierlijk virus dat in detail bestudeerd is op moleculair niveau eerste vectorontwikkelingen expressie recombinant eiwit
 zie ook Tabel voor eerste overzicht. Transiënte expressie - transiënte evaluatie van genexpressie / eiwitexpressie. testen van een grote reeks constructen in parallel - geen grootschalige. productie. analyse van regulatorische elementen (Box 12.1) (reportergenen !) Hoe afwezigheid van functionele ori - zolang plasmide stabiel - expressie. (’korte’ tijd = 1 of 2 dagen) ‘Runaway’ polyoma replicons – replicerend, maar celdood. Transiënt - episomaal. Ophoping van grote hoeveelheden DNA in korte tijd. Vb. Simian virus SV40 en murine (muis) polyomavirus. SV40, eerste dierlijk virus dat in detail bestudeerd is op moleculair niveau. eerste vectorontwikkelingen expressie recombinant eiwit.")
25
‘Runaway’ polyoma replicons – replicerend, maar celdood
SV40 circulair dsDNA, ~5 kb 2 transcriptie-eenheid : vroeg en laat meerdere producten door alternatieve splicing Vroeg: regulatorische eiwitten Laat: capside-eiwitten Regulatie: regulatorische sequenties tussen V/L regio Regulatorische regio : V/L promoter, enhancer, ori ; Vroeg : t en T tumorantigenen T vereist voor replicatie + onco- proteïne-eigenschappen

26
‘Runaway’ polyoma replicons - replicerend, maar celdood
SV40 – Virale vectoren Eerste vectoren, virale vectoren gentransfer via transductie Verplaatsing van vroege of late regio en functies voorzien in trans door co- introductie van een helpervirus Vereenvoudiging door COS-cellijn : gen van T antigen genomisch in trans voorzien bij vectoren met uitgewisselde vroege regio Nadeel : capaciteit viraal capside : max. ~ 2.5 kb te incorporeren SV40 ori op plasmiden Voordeel : geen grootterestrictie Enkel ori replicatie in COS cellijn ; ori + gen T-antigen permissieve cellen Transiënt hoge expressie – grote hoeveelheden recombinant eiwit Vb. pcDNA3.1 ‘runaway’-karakter veroorzaakt celdood !!! ontwikkeling van stabiele episomale vectoren door regulatie van T antigen expressie (conditionele promoter – temperatuursensitieve mutanten)
")
27
‘Runaway’ polyoma replicons – replicerend, maar celdood
BGH : “bovine growth hormone” sequenties pA : polyadenyleringssignaal PCMV = humane CMV (cytomegalovirus) promoter EK = enterokinase knipplaats voor verwijdering tags Hoge expressie in COS-cellen
 promoter. EK = enterokinase knipplaats voor verwijdering tags. Hoge expressie in COS-cellen.")
28
Human BK en BPV replicons – episomaal – structureel onstabiel
BKV Virussen die latente infecties veroorzaken Behoud als episomaal replicon (laag of matig aantal kopijen) Relatief laag kopijenaantal = replicon interfereert niet met gastheercelgroei Plasmiden met ‘latente’ ori (geen inpakking in capsiden) Voordeel t.o.v. integratieve vectoren: geen integratiespecifieke beïnvloeding van expressie (m.a.w. geen ‘positie’-effect) Humaan BK polyomavirus – infecteert meerdere celtypes – kopijenaantal ~500 kan opgedreven orden tot ~ 9000 (door toenemende concentratie ‘antibiotica’ waartegen een resistentiegen aanwezig is, bvb. neoR) BK T-antigen kan voorzien worden in trans
 Relatief laag kopijenaantal = replicon interfereert niet met gastheercelgroei. Plasmiden met ‘latente’ ori (geen inpakking in capsiden) Voordeel t.o.v. integratieve vectoren: geen integratiespecifieke beïnvloeding. van expressie (m.a.w. geen ‘positie’-effect) Humaan BK polyomavirus – infecteert meerdere celtypes – kopijenaantal ~500. kan opgedreven orden tot ~ 9000 (door toenemende concentratie ‘antibiotica’ waartegen een resistentiegen aanwezig is, bvb. neoR) BK T-antigen kan voorzien worden in trans.")
29
Human BK en BPV replicons – episomaal – structureel onstabiel
BKV ori T antigen - functies

30
Human BK en BPV replicons – episomaal – structureel onstabiel
BPV – Bovine papillomavirus Historisch - de eerste episomale vectoren (verre verwantschap met polyomavirussen) Kan muiscellen infecteren, zonder virus te produceren Kopijenaantal ~100 Ook T antigen (replicatie + oncogene transformatie gastheercel) (vroegste deel van infectie similair aan SV40 infectie, latere meer complex) Vroege functies op 69% fragment – BPV69T selectiemerker neo - selectie op geneticine Vb. expressie humaan beta-globine Nadeel : recombinatie - deleties - zekere vorm van instabiliteit
 Kan muiscellen infecteren, zonder virus te produceren. Kopijenaantal ~100. Ook T antigen (replicatie + oncogene transformatie gastheercel) (vroegste deel van infectie similair aan SV40 infectie, latere meer complex) Vroege functies op 69% fragment – BPV69T. selectiemerker neo - selectie op geneticine. Vb. expressie humaan beta-globine. Nadeel : recombinatie - deleties - zekere vorm van instabiliteit.")
31
Human BK en BPV replicons – episomaal – structureel onstabiel
BPV – Bovine papillomavirus

32
Epstein-Barr virus gebaseerde plasmiden – stabiele expressie
EBV : zeer stabiele replicatie in mens-cellen (en mammalia in het algemeen) EBV - herpesvirus - dsDNA ~170 kb mononucleosis (‘klierkoorts’) episomaal replicon, ~ 1000 kopijen onder natuurlijke omstandigheden : alleen infectie van lymfocyten, maar na transfectie ook in tal van andere primaatcellen te behouden Vereist voor episomale replicatie: latente oriP en trans-werkende regulator (EBNA1) EBV-gebaseerde expressievectoren – 2 tot 50 kopijen per cel Selectie : ‘neomycine’ of ‘hygromycine’ selectiemerkers - stabiele aanwezigheid Gebruik : episomale cDNA banken Introductie van oriP in YACs in vitro circularisatie introductie in humane cellen
 EBV - herpesvirus - dsDNA ~170 kb. mononucleosis (‘klierkoorts’) episomaal replicon, ~ 1000 kopijen. onder natuurlijke omstandigheden : alleen infectie van lymfocyten, maar na. transfectie ook in tal van andere primaatcellen te behouden. Vereist voor episomale replicatie: latente oriP en trans-werkende regulator (EBNA1) EBV-gebaseerde expressievectoren – 2 tot 50 kopijen per cel. Selectie : ‘neomycine’ of ‘hygromycine’ selectiemerkers - stabiele aanwezigheid. Gebruik : episomale cDNA banken. Introductie van oriP in YACs. in vitro circularisatie introductie in humane cellen.")
33
Epstein-Barr virus gebaseerde plasmiden – stabiele expressie

34
Bacteriële gentransfer – bacteriële vectoren
Transfer naar planten, m.b.v. Agrobacterium tumefaciens A. tumefaciens kan ook DNA introduceren in dierlijke cellen (2001) A. tumefaciens : transfer zonder invasie aanhechting en vorming pilus: conjugatieve transfer Bij dierlijke cellen : ‘bactofection’ o.a. Salmonella spp., Listeria monocytogenes en Shigella flexneri invasief opname in de cel, vervolgens lysis en vrijstelling van plasmide migreert naar nucleus ‘verzwakking’ (attenuation) nodig - zoniet vernietiging gastheercellen auxotrofe mutanten of stammen met induceerbare autolysis Toepassingen : o.a. gentransfer in vitro en in vivo - afleveren van recombinante DNA vaccins
 A. tumefaciens : transfer zonder invasie. aanhechting en vorming pilus: conjugatieve transfer. Bij dierlijke cellen : ‘bactofection’ o.a. Salmonella spp., Listeria monocytogenes en Shigella flexneri. invasief opname in de cel, vervolgens lysis en. vrijstelling van plasmide migreert naar nucleus. ‘verzwakking’ (attenuation) nodig - zoniet vernietiging gastheercellen. auxotrofe mutanten of stammen met induceerbare autolysis. Toepassingen : o.a. gentransfer in vitro en in vivo - afleveren van. recombinante DNA vaccins.")
35
Virussen als gentransfervectoren - transducerende vectoren
Transductie : horizontale DNA-transfer via mechanisme van virusinfectie Gentransfer in vitro (celculturen) en in vivo (gentherapie toepassingen) Insertie van een transgen of omwisseling van virale sequenties voor een transgen Insertie door ligatie of homologe recombinatie Helper-onafhankelijk : kan zich autonoom repliceren + propageren Helper-afhankelijk : virale elementen moeten in trans voorzien worden co-infectie met helpervirus ; helperplasmide voorzien of een ‘complementaire cellijn’ (ook ‘packaging’ cellijn) Vaak gewenst : afwezigheid van virale coderende sequenties enkel amplicons overhouden (cis-elementen) voordelen : niet cytotoxisch en grote inserten mogelijk Keuze virale vector : afhankelijk van te transformeren gastheer type expressie (stabiel – transiënt) hoeveelheid te verpakken DNA adenovirus – retrovirus – beperkte verpakkingscapaciteit (icosaëdrale hoofdjes) baculovirus – staafvormig – minder strikt
 en in vivo (gentherapie toepassingen) Insertie van een transgen of omwisseling van virale sequenties voor een transgen. Insertie door ligatie of homologe recombinatie. Helper-onafhankelijk : kan zich autonoom repliceren + propageren. Helper-afhankelijk : virale elementen moeten in trans voorzien worden. co-infectie met helpervirus ; helperplasmide voorzien of een ‘complementaire cellijn’ (ook ‘packaging’ cellijn) Vaak gewenst : afwezigheid van virale coderende sequenties enkel amplicons. overhouden (cis-elementen) voordelen : niet cytotoxisch en grote inserten mogelijk. Keuze virale vector : afhankelijk van te transformeren gastheer. type expressie (stabiel – transiënt) hoeveelheid te verpakken DNA. adenovirus – retrovirus – beperkte verpakkingscapaciteit. (icosaëdrale hoofdjes) baculovirus – staafvormig – minder strikt.")
36
Adenovirale vectoren voor kortstondige transiënte expressie
Adenovirus/afgeleide vectoren lineair, dsDNA genoom (~37 kb) 6 vroege transcriptie-eenheden (E) : virale replicatie 1 majeure late transcriptie-eenheid (MLT) : capside-eiwitten Veel gebruikt als gentransfer- & genexpressievector & attractief voor faagtherapie (efficiënte opname – neg./inflammatoire respons - immuunsysteem) Voordelen: stabiliteit, hoge capaciteit vreemd DNA, hoge titer (1011 pfu/ml), breed gastheerspectrum Transiënte expressie in delende cellen – geen efficiënte integratie Adenoviraal genoom
 6 vroege transcriptie-eenheden (E) : virale replicatie. 1 majeure late transcriptie-eenheid (MLT) : capside-eiwitten. Veel gebruikt als gentransfer- & genexpressievector & attractief voor faagtherapie. (efficiënte opname – neg./inflammatoire respons - immuunsysteem) Voordelen: stabiliteit, hoge capaciteit vreemd DNA, hoge titer (1011 pfu/ml), breed gastheerspectrum. Transiënte expressie in delende cellen – geen efficiënte integratie. Adenoviraal genoom.")
37
Adenovirale vectoren voor kortstondige transiënte expressie

38
Adenovirale vectoren:
- eerste generatie : replicatiedeficiënt : ‘E1 replacement vectors’ - kloneercapaciteit cfr. ‘uitgewisseld’ gedeelte ~7 kb - productie in de humane embryonale niercellijn 293 die de linker 11% van het adenovirus DNA bevatte complementatie van functies in trans - ook andere ‘E’-regio’s verwijderd eventueel grotere inserties (~10 kb capaciteit) - nadelen : virale expressie – cytotoxiciteit eventueel recombinatie met geïntegreerde comple- menterende sequenties - enkel amplicon overhouden - grote capaciteit (~ 37 kb) - minimaal cytotoxisch langere expressie - nadeel : geen cellijn die alle functies in trans voorziet verpakking vereist helpervirus risico van contaminatie
 - nadelen : virale expressie – cytotoxiciteit. eventueel recombinatie met geïntegreerde comple- menterende sequenties. - enkel amplicon overhouden. - grote capaciteit (~ 37 kb) - minimaal cytotoxisch langere expressie. - nadeel : geen cellijn die alle functies in trans voorziet. verpakking vereist helpervirus risico van contaminatie.")
39
Vectorafgeleiden van het adeno-geassocieerd virus - integratief
Adeno-associated virus (AAV) AAV : contaminant in adenoviraal preparaat ssDNA virus - Parvoviridae AAV-genoom : ~5 kb rep (replicatie) en cap (capside) genen geflankeerd door 145 nt omgekeerde herhalingen
 AAV : contaminant in adenoviraal preparaat. ssDNA virus - Parvoviridae. AAV-genoom : ~5 kb. rep (replicatie) en cap (capside) genen geflankeerd door 145 nt. omgekeerde herhalingen.")
40
Van nature replicatiedeficiënt - vereist ander virus (e. g
Van nature replicatiedeficiënt - vereist ander virus (e.g. adenovirus, herpesvirus) voor lytische infectiecyclus Zonder ‘helper’ : integratie in het genoom (constante locus - chromosoom 19) - latent provirus ‘rescue’ door o.a. een adenovirus-infectie Noodzaak van helperinfectie = controle over virusreplicatie – veiligheid bij gentherapietoepassing (samen met specifieke integratie) Vectorontwikkeling : rep proteïnen interfereren met endogene promoters cytotoxisch effect deletie rep en cap genen LTR enige noodzakelijke elementen voor replicatie, transcriptie, provirale integratie (eventueel niet plaatsspecifiek in afwezigheid van rep-eiwitten) en ‘rescue’. Transfectie van AAV-afgeleid construct : LTR + transgen (in plasmidevector) + AAV-functies (helperplasmide) + transfectie-gebaseerd adenoviraal helperplasmide (virusvorming) + affiniteitschromatografie voor opzuivering AAV-partikels – zoniet contaminatie met adenoviruspartikels In principe beperkte capaciteit verpakking vreemd DNA - aflevering van beperkte hoeveelheid DNA omzeilen via co-introductie van 2 segmenten op 2 vectoren vectorconcatemerisatie in doelwitcel
 voor lytische infectiecyclus. Zonder ‘helper’ : integratie in het genoom (constante locus - chromosoom 19) - latent provirus ‘rescue’ door o.a. een adenovirus-infectie. Noodzaak van helperinfectie = controle over virusreplicatie – veiligheid bij. gentherapietoepassing (samen met specifieke integratie) Vectorontwikkeling : rep proteïnen interfereren met endogene promoters cytotoxisch effect. deletie rep en cap genen LTR enige noodzakelijke elementen voor. replicatie, transcriptie, provirale integratie (eventueel niet plaatsspecifiek in. afwezigheid van rep-eiwitten) en ‘rescue’. Transfectie van AAV-afgeleid construct : LTR + transgen (in plasmidevector) + AAV-functies (helperplasmide) + transfectie-gebaseerd adenoviraal helperplasmide (virusvorming) + affiniteitschromatografie voor opzuivering AAV-partikels – zoniet. contaminatie met adenoviruspartikels. In principe beperkte capaciteit verpakking vreemd DNA - aflevering van. beperkte hoeveelheid DNA omzeilen via co-introductie van 2. segmenten op 2 vectoren vectorconcatemerisatie in doelwitcel.")
41
Baculovirus-afgeleide vectoren: hoog expressieniveau in insectcellijnen
staafvormig capside groot dsDNA genoom gastheer: insekten bepaalde baculovirussen : polyhedrosis virussen – productie van ‘nuclear occlusion bodies’ virions omgeven door eiwitlaag (polyhedrine) – extra bescherming polyhedrine productie – hoog – sterke promoter polyhedrine niet essentieel polyhedrine-gen uit te wisselen voor een transgen = polyhedrine ‘replacement’ vectoren (hoog expressieniveau mg/106 cellen) Meest toegepast : ‘Autographa californica multiple nuclear polyhedrosis virus’ (AcMNPV) eiwitexpressie in cellijnen (bvb. Sf9, Sf21) ‘Bombyx mori nuclear polyhedrosis virus’ (BmNPV) infectie zijderups – recombinante eiwitexpressie in organisme
 – extra bescherming. polyhedrine productie – hoog – sterke promoter. polyhedrine niet essentieel. polyhedrine-gen uit te wisselen voor een transgen. = polyhedrine ‘replacement’ vectoren. (hoog expressieniveau - 1 mg/106 cellen) Meest toegepast : ‘Autographa californica multiple nuclear polyhedrosis virus’ (AcMNPV) eiwitexpressie in cellijnen (bvb. Sf9, Sf21) ‘Bombyx mori nuclear polyhedrosis virus’ (BmNPV) infectie zijderups – recombinante eiwitexpressie in organisme.")
42
Baculovirus-afgeleide vectoren: hoog expressieniveau in insectcellijnen
Endocytosis van baculovirus

43
Glycosylatie : - mogelijk - echter : glycosylatiereactieweg verschillend bij insecten en zoogdieren mogelijk : onjuiste glycosylatie immunogeen recombinant eiwit - oplossing : cellijnen die zoogdiereigen glycosylatie uitvoeren of co-expressie van geschikte glycosylatie-enzymen - vb. expressie van plasminogeenactivator (gegalactosyleerd) In de Sf9-cellijn die 1,4-galactosyltransferase tot expressie brengt onder een vroege baculoviruspromoter. Zodra infectie met recombinant baculovirus (met plasminogeenactivatorgen) ook mogelijkheid van correcte glycosylatie ‘replacement vectors’ uitwisseling van polyhedrine-gen : screening van recombinanten : * Wild-type virus productie occlusion bodies opaque plaques Recombinante geen occlusion bodies heldere plaques * E. coli lacZ gen in leesraam blauw-wit onderscheid * ‘Green fluorescent eiwit’ (GFP) in leesraam
 In de Sf9-cellijn die 1,4-galactosyltransferase tot expressie brengt. onder een vroege baculoviruspromoter. Zodra infectie met. recombinant baculovirus (met plasminogeenactivatorgen) ook. mogelijkheid van correcte glycosylatie. ‘replacement vectors’ uitwisseling van polyhedrine-gen : screening van. recombinanten : * Wild-type virus productie occlusion bodies opaque plaques. Recombinante geen occlusion bodies heldere plaques. * E. coli lacZ gen in leesraam blauw-wit onderscheid. * ‘Green fluorescent eiwit’ (GFP) in leesraam.")
44
Klonering door homologe recombinatie tussen polyhedrine-gedeleteerd
viraal DNA en vector met transgen en homologe regio’s * nadeel bij beperkte deletie : lage frequentie recombinanten * ontw. : grote deletie in genoom + gelineariseerd grote deletie enkel te herstellen door homologe recombinatie met vector vector overspant de deletie + flankerende regio’s voor homologe recombinatie leefbaar, recombinant genoom tot 90% recombinanten

45
‘polyhedrine replacement vectors’
Opmerking bij illustratie meer succesvolle systemen met grotere deletie dan enkel het polyhedrine-gen

46
Baculovirus-afgeleide vectoren: hoog expressieniveau in insectcellijnen

47
Herpesvirus vectoren – latent – mogelijk lange termijn transgenexpressie
Herpesvirussen – groot dsDNA – o.a. EBV, HSV, Varicella zoster HSV-I gebaseerde transducerende vectoren na transfer door transfectie – compatiebel met vele cellijnen evenwel: via transductie – voor gentherapie – neurotopisch = gericht naar zenuwcellen groot virus – transgenklonering via homologe recombinatie in getransfecteerde cellen Alternatief: plasmide-gebaseerde ampliconvectoren – enkel cis-elementen (repl. & verpak- king) + verpakkingssystemen voorzien in trans gentransfer in vivo naar neuronen
 + verpakkingssystemen voorzien in trans. gentransfer in vivo naar neuronen.")
48
Retrovirale vectoren – efficiënte genomische integratie
Retrovirussen: RNA-virussen die repliceren via DNA-intermediair Integratie in genoom gastheercel Vectorontwikkeling – belangrijke kenmerken: - sommige acuut oncogeen – replicatie-deficiënt - geen gastheerafdoding – continue productie - vaak sterke promoters – hoge eiwitexpressie (mogelijk induceerbaar) - sommige breed celspectrum - geschikt voor vectorontwikkeling door beperkte genoomgrootte (in vitro-manipulaties eenvoudig) – hoge titers – efficiënte infectie Nadelen: velen – enkel efficiënte infectie van delende cellen beperkt gentherapie- toepassingen Lentiviridae (o.a. HIV) wel mogelijkheid infectie niet-delende cellen
 - sommige breed celspectrum. - geschikt voor vectorontwikkeling door beperkte genoomgrootte. (in vitro-manipulaties eenvoudig) – hoge titers – efficiënte infectie. Nadelen: velen – enkel efficiënte infectie van delende cellen beperkt gentherapie- toepassingen. Lentiviridae (o.a. HIV) wel mogelijkheid infectie niet-delende cellen.")
49
Oncoretrovirussen - genoom
Boven: geïntegreerd provirus - LTR Onder : Verpakt RNA PB = primer binding sites – virale replicatie

50
Geïntegreerd genoom bevat gag, pol, env genen
gag structurele eiwitten pol reverse transcriptase env enveloppe-eiwitten Viraal RNA afgeschreven van promoter in linker LTR en stopt na polyadenylatiesignaal in rechter LTR. Ook capping. Gag-pol fusieproteïne processing tot meerdere polypeptiden Deel RNA splicing gag-pol en translatie env Twee kopijen RNA genoom verpakt + reverse transcriptase & integrase

Verwante presentaties
>")










.>")







 enzymen.>")