Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Klonering van grote DNA segmenten : BAC, PAC, YAC vectoren.
cfr. o.a. Primrose & Twyman : hfdst. 5 Cosmiden : inserts kbp : selectie door inpakrestrictie in faag l partikels (zie Gentechnologie) Zoektocht naar fagen met grotere inpakmogelijkheden (o.a. T4) succesvol met faag P1 (E. coli getemperde faag, extensief gebruikt voor algemene transductie) Gebruik van F-plasmide als replicon, samen met elektroporatie als DNA-transformatietechniek leidde in 1992 tot BAC vectoren. ('bacterial artificial chromosome') Combinatie van P1 en BAC eigenschappen leidde in 1994 tot PAC vectoren. ('P1-derived artificial chromosome'). Met BAC en PAC kunnen DNA segmenten tot 300 kb gekloneerd worden, hoewel (gezien er geen grootte-selectie is) vaak inserts van kb (of zelfs minder) bekomen worden. DNA isolatie, en intactheid, spelen uiteraard een belangrijke rol. Het concept (en terminologie) "artificial chromosome" was reeds in 1987 ontwikkeld in S. cerevisiae met de YAC vectoren (bij de studie van stabiliteit in gist bij toevoeging van telomeersequenties aan YCp vector DNA).
 Zoektocht naar fagen met grotere inpakmogelijkheden (o.a. T4) succesvol met faag P1. (E. coli getemperde faag, extensief gebruikt voor algemene transductie) Gebruik van F-plasmide als replicon, samen met elektroporatie als. DNA-transformatietechniek leidde in 1992 tot BAC vectoren. ( bacterial artificial chromosome ) Combinatie van P1 en BAC eigenschappen leidde in 1994 tot PAC vectoren. ( P1-derived artificial chromosome ). Met BAC en PAC kunnen DNA segmenten tot 300 kb gekloneerd worden, hoewel (gezien er geen grootte-selectie is) vaak inserts van kb. (of zelfs minder) bekomen worden. DNA isolatie, en intactheid, spelen uiteraard een belangrijke rol. Het concept (en terminologie) artificial chromosome was reeds in ontwikkeld in S. cerevisiae met de YAC vectoren (bij de studie van stabiliteit in gist bij toevoeging van telomeersequenties aan YCp vector DNA).")
2
E. coli bacteriofaag P1 getemperde faag van E. coli.
medieert algemene transductie. het genoom is circulair gepermuteerd en terminaal redundant (10 kb). het unieke (niet-gerepeteerde) DNA is ongeveer 90 kb. bij lysogenie integreert het faag DNA niet in het chromosoom maar blijft als extrachromosomaal DNA in laag aantal kopijen in de cel ("als plasmide"). bij inductie tot lytische ontwikkeling fungeert een lytisch replicon tot hoog aantal kopijen. in het faaggenoom volgt na pac een loxP plaats : bij concatenatie volgen opeenvolgende loxP's elkaar op per ~90 kb. Pacase splitst per ~110 kb, zodat het eerste product 2 loxP sequenties heeft 2 loxP-sequenties : worden herkend door het product van het cre gen (Cre recombinase) circularisatie van (sommige) DNA moleculen in E. coli in 1990 ontwikkeld tot een vectorsysteem waarin ~100 kb insereerbaar is totale capaciteit inpakbaar DNA: ~110 kb pac (‘packaging site’) sequentie : startplaats voor verpakking essentieel element, splitsing door Pacase kan ook in vitro gebruikt worden
. het unieke (niet-gerepeteerde) DNA is ongeveer 90 kb. bij lysogenie integreert het faag DNA niet in het chromosoom maar blijft als. extrachromosomaal DNA in laag aantal kopijen in de cel ( als plasmide ). bij inductie tot lytische ontwikkeling fungeert een lytisch replicon tot hoog aantal. kopijen. in het faaggenoom volgt na pac een loxP plaats : bij concatenatie volgen. opeenvolgende loxP s elkaar op per ~90 kb. Pacase splitst per ~110 kb, zodat het eerste product 2 loxP sequenties heeft. 2 loxP-sequenties : worden herkend door het product van het cre gen (Cre recombinase) circularisatie van (sommige) DNA moleculen in E. coli. in 1990 ontwikkeld tot een vectorsysteem waarin ~100 kb insereerbaar is. totale capaciteit inpakbaar DNA: ~110 kb. pac (‘packaging site’) sequentie : startplaats voor verpakking. essentieel element, splitsing door Pacase. kan ook in vitro gebruikt worden.")
3
Het concatemeer DNA wordt gesplitst op opeenvolgende 100 kb posities (vanaf de pac).
loxP plaatsen liggen ongeveer 90 kb uit elkaar (het unieke DNA in het P1 genoom). Er worden ongeveer 4 faagkoppen gevuld per concatemeer DNA, maar slechts één heeft twee loxP’s, en kan dus cycliseren in een recA E. coli mutant. (Cre is faaggekodeerd.)
. Er worden ongeveer 4 faagkoppen gevuld per concatemeer DNA, maar slechts één heeft twee loxP’s, en kan dus cycliseren in een recA E. coli mutant. (Cre is faaggekodeerd.)")
4
Vector-DNA wordt bereid als twee armen, één met pac + loxP, de andere met
de replicatiefuncties, resistentiemerker en loxP. In een trimoleculaire ligatiereactie wordt insert-DNA ingebouwd. Transductie naar een E. coli die het Cre-recombinase tot expressie brengt, zodat het verpakte DNA kan circulariseren Klonen met laag-kopijaantal – selectiemerker/Kanamycine resistentie Eventueel hoger kopijenaantal via het P1 lytisch replicon Inserts tussen 70 en 100 kb Voorbeelden: P1 vector Ad10 (1990), pAD10sacBII Het inpakmechanisme gebeurt via een "headful" mechanisme, zodat voldoende "extra" DNA in de lange arm als "stuffer" aanwezig moet zijn. Oorspronkelijk werd daarvoor adenovirus DNA gebruikt, hetgeen de nomenclatuur van de eerste (en ook latere) vectoren verklaart. In latere vectoren werd vooral het sacB ingevoerd voor positieve selectie van recombinanten, evenals faag RNA-promotoren rond de insertieplaats om makkelijk RNA sondes te kunnen maken.
, pAD10sacBII. Het inpakmechanisme gebeurt via een headful mechanisme, zodat. voldoende extra DNA in de lange arm als stuffer aanwezig moet. zijn. Oorspronkelijk werd daarvoor adenovirus DNA gebruikt, hetgeen. de nomenclatuur van de eerste (en ook latere) vectoren verklaart. In latere vectoren werd vooral het sacB ingevoerd voor positieve. selectie van recombinanten, evenals faag RNA-promotoren rond de. insertieplaats om makkelijk RNA sondes te kunnen maken.")
5
Faag P1 vectorsysteem

6
Faag P1 vector pAd10SacBII

7
Faag P1 vector pAd10SacBII
Kenmerken 2 loxP sequenties pac sequentie: verpakking in P1 faagpartikels kanr : kanamycineresistentiegen P1 plasmide-replicon: circulair DNA aan ~1 kopij/cel P1 lytisch replicon (‘lytisch replicon’) : onder controle van de lac promoter (induceerbaar met IPTG) : plasmide-amplificatie vóór DNA-isolatie sacB: positieve selectiemerker voor klonen met insert-DNA (BamHI-site) met faag SP6 en T7 promoter
 : onder controle van de lac promoter. (induceerbaar met IPTG) : plasmide-amplificatie vóór DNA-isolatie. sacB: positieve selectiemerker voor klonen met insert-DNA (BamHI-site) met faag SP6 en T7 promoter.")
8
BAC vectoren : bacterieel artificieel chromosoom (1992)
vectoren ontwikkeld op basis van het E. coli F plasmide : 1 of 2 kopijen per cel replicatie (unidirectioneel) vereist oriS en repE RepE : helicase (bevordert DNA replicatie) parA, parB en parC voor correcte partitie over de dochtercellen (stabiel behoud van het plasmide) antibioticumresistentiemerker nodig voor selectie insertie van DNA door in vitro ligatie (later, eventueel door recombinatie) er is geen selectie op insertgrootte transformatie door electroporatie ; geen verpakkingsextracten vereist (evenmin mogelijk) bij voorkeur gebruik van recA- E. coli waardcel structureel-stabiel behoud van mens- en plant-DNA (> 100 generaties) nieuwere BAC-vectoren : met lacZa of sacB gen voor herkenning of positieve selectie van recombinanten unieke splitsbaarheid van grote DNA's vergemakkelijkt door het voorzien van cosN, loxP, SceI, NotI, SfiI, ... T7 en SP6 promotoren voor aanmaak van RNA sondes zeer efficient voor genoomkartering en isolatie van operons, enz. Manipuleerbaarheid van het gekloond DNA is moeilijker gezien de grootte (unieke knipplaatsen?).
 vereist oriS en repE. RepE : helicase (bevordert DNA replicatie) parA, parB en parC voor correcte partitie over de dochtercellen. (stabiel behoud van het plasmide) antibioticumresistentiemerker nodig voor selectie. insertie van DNA door in vitro ligatie (later, eventueel door recombinatie) er is geen selectie op insertgrootte. transformatie door electroporatie ; geen verpakkingsextracten vereist. (evenmin mogelijk) bij voorkeur gebruik van recA- E. coli waardcel. structureel-stabiel behoud van mens- en plant-DNA (> 100 generaties) nieuwere BAC-vectoren : met lacZa of sacB gen voor herkenning of positieve. selectie van recombinanten. unieke splitsbaarheid van grote DNA s vergemakkelijkt door het voorzien van cosN, loxP, SceI, NotI, SfiI, ... T7 en SP6 promotoren voor aanmaak van RNA sondes. zeer efficient voor genoomkartering en isolatie van operons, enz. Manipuleerbaarheid. van het gekloond DNA is moeilijker gezien de grootte (unieke knipplaatsen ).")
9
Voorbeeld : pBeloBACII

10
PAC vectoren : faag P1-afgeleid artificieel chromosoom 1994
Combinatie van elementen uit de BAC en faag P1 vectorsystemen replicatie zoals P1-vectoren, maar ontdaan van verpakkingssignalen sacB, positieve selectiemerker P1 plasmide en lytisch replicon geen verpakking in P1 faagpartikels geen cre-loxP systeem voor circularisatie – circularisatie bij in vitro ligatie introductie door elektroporatie Voorbeeld: pCYPAC1 insertgroottes bij klonering in BACs of PACs zijn vergelijkbaar ; eventuele geobserveerde verschillen hebben allicht eerder met DNA bereiding te maken dan met inherente karakteristieken. Inserties tot 300 kb bekomen ; DNA-banken hebben meestal een gemiddelde grootte tussen 80 en 120 kb.

11
Voorbeeld : pCYPAC1

12
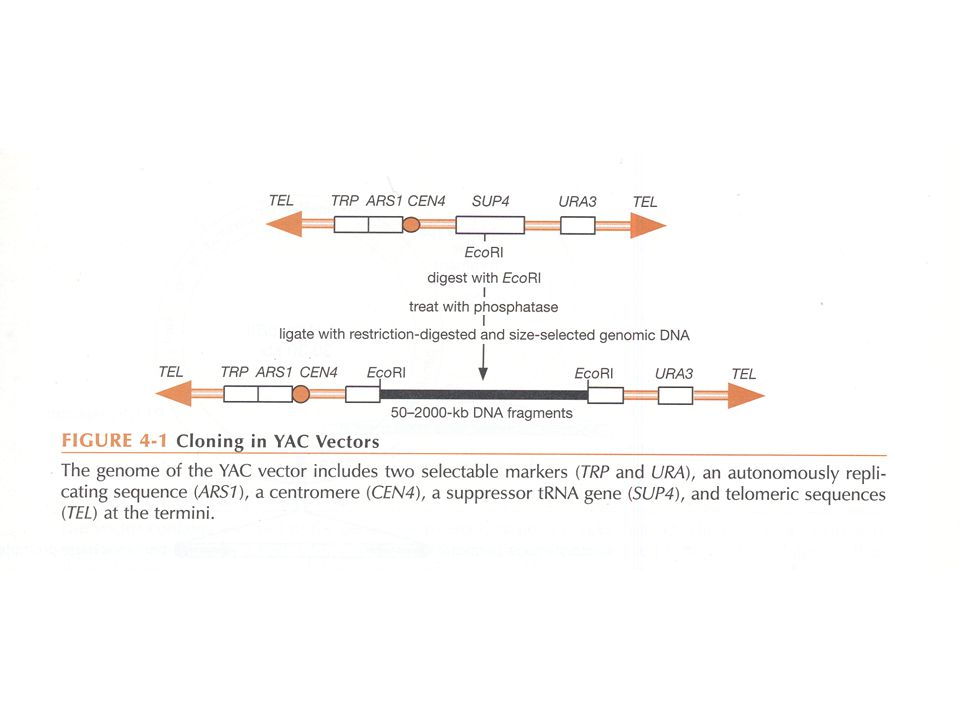
YAC-vectoren : ‘yeast’ artificieel chromosoom (gist) 1987
"Klassieke" YACs : lineair DNA : architectuur zoals gist chromosoom in 1982 werd aangetoond dat in gist een extra DNA molecule lineair behouden kon blijven mits toevoeging van telomeer sequenties in een YRp; vereiste voor stabiliteit : minimum kbp lengte in 1987 werd een kloneringsprocedure uitgewerkt : YAC vectoren recombinante YAC’s door ligatie van grote DNA fragmenten aan twee “armen” introductie in gist door transformatie op iedere arm : een selecteerbare merker, een telomeer sequentie - één arm bevat een centromeer, ARS (replicatie in gist), en ori (voor E. coli) - er is tevens een selectiemerker voor E. coli - om aanmaak van de vector in E. coli mogelijk te maken is er een verbindings-DNA tussen de telomeren, uitsplitsbaar met BamHI de twee vectorarmen worden dus aangemaakt door splitsing met EcoRI + BamHI selectie van gisttransformanten op selectieve agar (TRP, URA3 in de voorbeelden) recombinanten kunnen herkend worden door een wit => rood omkleuring : gebaseerd op een suppressor tRNA gen (SUP4) dat een 'ochre' nonsense mutatie (UAA) in het ade2 gen onderdrukt (waardoor witte kleur). minimum-grootte nodig voor normale “segregatie” van YACs (<20 kbp)
, en ori (voor E. coli) - er is tevens een selectiemerker voor E. coli. - om aanmaak van de vector in E. coli mogelijk te maken is er een. verbindings-DNA tussen de telomeren, uitsplitsbaar met BamHI. de twee vectorarmen worden dus aangemaakt door splitsing met EcoRI + BamHI. selectie van gisttransformanten op selectieve agar (TRP, URA3 in de voorbeelden) recombinanten kunnen herkend worden door een wit => rood omkleuring : gebaseerd. op een suppressor tRNA gen (SUP4) dat een ochre nonsense mutatie (UAA) in het ade2 gen onderdrukt (waardoor witte kleur). minimum-grootte nodig voor normale segregatie van YACs (<20 kbp)")
13
yeast artificial chromosome (YAC)
Lineaire YAC (met telomeersequenties) versus “Circulaire” YAC “gecirculariseerde” YAC (“BAC”) – geen telomeer- Seq.
 versus. Circulaire YAC. gecirculariseerde YAC. ( BAC ) – geen telomeer- Seq.")
15
Voorzorgen : bij bewerkingen : mogelijk probleem bij manipulaties van grote DNAs: intact houden ! kleine fragmenten verwijderen door PFGE fractionatie voor klonering polyamines toevoegen (inhibeert DNA afbraak) => de inserts van de eerste YAC-DNA banken waren niet groter dan 100 kb Toch zijn er uiteindelijk (na optimalisatie) YAC-banken gemaakt van het menselijk genoom met inserts van gemiddeld 810 kb, zelfs sommige tot 1800 kb. mogelijkheid om grote intacte genen te isoleren (die bij hogere eukaryoten meer dan 1Mb kunnen zijn) Mogelijke problemen met klassieke (lineaire) YACs: 20-60% zijn chimere YACs : verschillende genomische regio’s gekloneerd in één YAC (co-ligatie vóór transformatie of recombinatie in gist) (lijkt vooral bij protoplast-stap bij transformatie op te treden) => beter intacte cellen gebruiken instabiliteit : verlies van YACs en deletie van interne regio’s in de YACs (gedurende transformatie en groei/deling) oplossing: recombinatiedeficiënte giststammen gebruiken ; echter daardoor trage groei + inefficiënte transformatie (behalve met het rad54-3 allel) Lage opbrengst + moeilijk te scheiden van gastheer genomisch DNA (15 Mb) PFGE
 => de inserts van de eerste YAC-DNA banken waren niet groter dan 100 kb. Toch zijn er uiteindelijk (na optimalisatie) YAC-banken gemaakt van het menselijk. genoom met inserts van gemiddeld 810 kb, zelfs sommige tot 1800 kb. mogelijkheid om grote intacte genen te isoleren. (die bij hogere eukaryoten meer dan 1Mb kunnen zijn) Mogelijke problemen met klassieke (lineaire) YACs: 20-60% zijn chimere YACs : verschillende genomische regio’s gekloneerd in één YAC. (co-ligatie vóór transformatie of recombinatie in gist) (lijkt vooral bij protoplast-stap bij transformatie op te treden) => beter intacte cellen gebruiken. instabiliteit : verlies van YACs en deletie van interne regio’s in de YACs (gedurende. transformatie en groei/deling) oplossing: recombinatiedeficiënte. giststammen gebruiken ; echter daardoor trage groei + inefficiënte. transformatie (behalve met het rad54-3 allel) Lage opbrengst + moeilijk te scheiden van gastheer genomisch DNA (15 Mb) PFGE.")
16
OPLOSSING : Circulaire YACs :
makkelijk en snel naar E. coli overbrengbaar makkelijk isoleerbaar via alkalische DNA bereiding (circulair cccDNA denatureert/renatureert efficienter) circulaire YACs die ook te manipuleren zijn in E. coli als BACs een "gespecialiseerde BAC" waarop - een gist centromeersequentie (CEN6 in de figuur), - een G418R selectiemerker, en HIS - alle BAC elementen voor behoud in E. coli - twee homologe sequenties die respectievelijk aan deze en gene zijde van van de kloneerplaats in de klassieke YAC-vector liggen (dit worden de plaatsen waar door recombinatie het insert wordt overgeplaatst) ("hooks" genoemd : de hooks zijn gescheiden door een unieke SalI knipplaats) linearisatie van deze “BAC” met SalI, gevolgd door recombinatie in gist en het ontstaan van circulaire YAC (repliceerbaar in gist door CEN6 + ARS) Voordelen: nu gemakkelijk te scheiden van de lineaire chromosomen : - alkalische hydrolyse, minder shearing-gevoelig. - grotere structurele stabiliteit ( kb inserts vergelijkbare stabiliteit met BAC-kloons) - grotere inserten : tot 40% chimeren, maar niettemin structureel stabiel => bestudeerbaar - elektroporeerbaar naar E. coli (tot kb)
 circulaire YACs die ook te manipuleren zijn in E. coli als BACs. een gespecialiseerde BAC waarop. - een gist centromeersequentie (CEN6 in de figuur), - een G418R selectiemerker, en HIS. - alle BAC elementen voor behoud in E. coli. - twee homologe sequenties die respectievelijk aan deze en gene zijde van. van de kloneerplaats in de klassieke YAC-vector liggen. (dit worden de plaatsen waar door recombinatie het insert wordt overgeplaatst) ( hooks genoemd : de hooks zijn gescheiden door een unieke SalI knipplaats) linearisatie van deze BAC met SalI, gevolgd door recombinatie in gist en het ontstaan. van circulaire YAC (repliceerbaar in gist door CEN6 + ARS) Voordelen: nu gemakkelijk te scheiden van de lineaire chromosomen : - alkalische hydrolyse, minder shearing-gevoelig. - grotere structurele stabiliteit ( kb inserts vergelijkbare stabiliteit met BAC-kloons) - grotere inserten : tot 40% chimeren, maar niettemin structureel stabiel => bestudeerbaar. - elektroporeerbaar naar E. coli (tot kb)")
17
Circulaire YAC Recombinatie tussen YAC met genomisch insert en de “BAC” met functionele gistelementen (CEN6 sequentie – centromeer, selectiemerkers – geen ARS in “BAC” dus de “BAC” repliceert niet in gist)
")
18
TAR : (transformation-associated recombination)
Similair zoals hierboven, maar nu als doelwit een sequentie die tussen de promotor van ADH1 (S. pombe alcohol dehydrogenase) gen ligt en het URA3 gen (zonder eigen promotor). Deze promotor laat tot 130 bp toe tussen de TATA box en de transcriptie-initiatieplaats. Hier kunnen dus twee 'hooks' van 60 bp liggen. NB. ade2 is een mutant allel van fosforibosylamino-imidazole carboxylase. De mutatie is een nonsense mutatie (ochre) die door Sup4 gesuppresseerd kan worden. Transformanten met de parentale vector (actief Sup4) groeien tot witte kolonies op een medium van zuur-gehydrolyseerd caseïne. Bij recombinanten (Sup4 geïnactiveerd) is het enzym niet aanwezig en accumuleert fosforibosylglycinamide in de cellen, tot rood-roze kolonies.
 gen ligt en. het URA3 gen (zonder eigen promotor). Deze promotor laat tot 130 bp toe tussen de TATA box en de. transcriptie-initiatieplaats. Hier kunnen dus twee hooks van. 60 bp liggen. NB. ade2 is een mutant allel van fosforibosylamino-imidazole carboxylase. De mutatie is een nonsense mutatie (ochre) die door Sup4 gesuppresseerd kan worden. Transformanten met de parentale vector (actief Sup4) groeien tot witte kolonies op een. medium van zuur-gehydrolyseerd caseïne. Bij recombinanten (Sup4 geïnactiveerd) is het enzym niet aanwezig en accumuleert. fosforibosylglycinamide in de cellen, tot rood-roze kolonies.")
19
Overzicht : kloneren van grote fragmenten
Kloneren van grote fragmenten : cosmiden, P1, PAC, BAC, YAC - overzicht Vector Capaciteit Replicon Gastheer Kopijenaantal Introductie (kb) Cosmide ColE E. coli hoog transductie P P E. coli (amplificeerbaar) transductie PAC P E. coli elektroporatie BAC F E. coli elektroporatie YAC ARS gist transformatie P1, cosmiden: insertiegrootte beperkt door verpakkingsmogelijkheid in faagpartikels BAC, PACs: minder chimerisatie, eenvoudige bankconstructie, eenvoudige DNA-manipulaties – klonering/insertie YACs: chimerisatie/stabiliteit Belang : - kartering en analyse van complexe genomen ; isolatie operons - beter zicht op overlappende klonen – naburige sequenties in grote genomen - opstellen van fysische kaarten - inzoemen op bepaalde genomische regio’s/operons (sequentieanalyse & hybridisatie)
 Cosmide ColE1 E. coli hoog transductie. P P1 E. coli 1 (amplificeerbaar) transductie. PAC P1 E. coli 1 elektroporatie. BAC F E. coli 1 elektroporatie. YAC ARS gist 1 transformatie. P1, cosmiden: insertiegrootte beperkt door verpakkingsmogelijkheid in faagpartikels. BAC, PACs: minder chimerisatie, eenvoudige bankconstructie, eenvoudige. DNA-manipulaties – klonering/insertie. YACs: chimerisatie/stabiliteit. Belang : - kartering en analyse van complexe genomen ; isolatie operons. - beter zicht op overlappende klonen – naburige sequenties in grote genomen. - opstellen van fysische kaarten. - inzoemen op bepaalde genomische regio’s/operons (sequentieanalyse & hybridisatie)")
20
vervolgt…

21
…vervolg

Verwante presentaties




















 tot hs 30>")