Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Enzymen en enzymkinetiek
IDB HC3 en HC4 Enzymen en enzymkinetiek Michaelis Menten Inhibitie van enzymen Ivo Horn

2
Te bestuderen Wilson and Walker, Principles and techniques of biochemistry and molecular biology: blz Campbell and Farrell, 7e editie: H6, optioneel: Campbell and Reece, 9e editie: H8: 8.4

3
Biologische katalyse Katalyse is de belangrijkste functie van eiwitten
Katalyse: versnellen/efficienter laten verlopen van een reactie Biologische katalytische eiwitten: enzymen Verhogen de snelheid niet-enzymen komen tot 104 Er zijn enkele niet-eiwit katalytische biomoleculen: ribozymen. Dit zijn katalytische RNA’s

4
Biologische wasmiddelen
Bevatten enzymen Unilever research Hittebestendig maken van eiwitten! Normale enzymen werken optimaal bij 37 graden Celsius

5
Een enzym betrokken bij ziekten
Het GTPase Ras is een enzym Zet GTP in GDP om Verandert daarbij van conformatie/vorm De GTP –gebonden vorm is aktief en geeft signalen door in de cel In bijvoorbeeld bepaalde tumoren is Ras gemuteerd en constant in de aktieve conformatie

6
Acetylcholine esterase (ACE)
Bij Alzheimer: het enzym acetylcholine esterase wordt geremd waardoor afbraak van acetylcholine tegen wordt gegaan

7
Functies van eiwitten:
Enzymen: versnellen reacties Structurele eiwitten: collageen (in bot) en keratine (in haar) Opslag eiwitten: albumine (in bloed) en caseïne (in melk) Transport eiwitten: Hb (O2 in bloed), albumine (bili in bloed) Hormonale eiwitten: insuline, glucagon Receptor eiwitten Contractiele eiwitten in spierweefsel Defensieve eiwitten: antilichamen
 en keratine (in haar) Opslag eiwitten: albumine (in bloed) en caseïne (in melk) Transport eiwitten: Hb (O2 in bloed), albumine (bili in bloed) Hormonale eiwitten: insuline, glucagon. Receptor eiwitten. Contractiele eiwitten in spierweefsel. Defensieve eiwitten: antilichamen.")
8
PKMζ

9
Indeling van enzymen: Transferases (verplaatsen groepen op eiwitten)
Hydrolases (maken water vrij) Ligases (“lijmen” DNA stukken aan elkaar) Kinases (fosforyleren eiwitten) GTPases (zetten GTP om in GDP) Fosforylases (verwijderen fosfaatgroepen) etc
 Ligases ( lijmen DNA stukken aan elkaar) Kinases (fosforyleren eiwitten) GTPases (zetten GTP om in GDP) Fosforylases (verwijderen fosfaatgroepen) etc.")
10
geheugenfunctie PKMζ is betrokken bij geheugenvorming
Muizen kregen saccharine toegediend Misselijkheid werd opgewekt direct daarna Reactie later op saccharine: misselijkheid (geheugen!) Injectie ZIP in de cortex, geen misselijkheid. Remming enzym PKMζ
 Injectie ZIP in de cortex, geen misselijkheid. Remming enzym PKMζ.")
11
Voorbeeld groep 1 CH3CH2OH + NAD+ CH3CHO + NADH+ H+ ethanol ethanal Alcoholdehydrogenase (alcohol:NAD oxidoreductase) Nummer (EC-rules) 1.Hoofdgroep 1.CH-OH is donor 1.NAD+ of NADP+ is acceptor 1.Eerst gevonden enzym in deze groep Verwijdert protonen en vormt daardoor ethanal uit ethanol
 Nummer (EC-rules) 1.Hoofdgroep 1.CH-OH is donor 1.NAD+ of NADP+ is acceptor 1.Eerst gevonden enzym in deze groep Verwijdert protonen en vormt daardoor ethanal uit ethanol")
12
Overzicht Katalytisch vermogen,specificiteit, regulatie
Introductie enzymkinetiek Kinetiek van enzym-gekatalyseerde reacties Enzyminhibitie Ribozymen

13
Enzymen Enzymen laten cellen beschikken over het vermogen om reacties zo snel te laten verlopen als nodig is voor de cel Enzymen zijn de stoffen die gerelateerd zijn aan een metabolische functie

14
Gekatalyseerde reacties vergen minder activatie energie

15
De DG waarde Negatief: exergone reactie. Spontaan verlopende reactie; er komt energie vrij Positief: endergone reactie. Reactie vraagt energie om te verlopen. Energetisch dus ongunstig Nul: exergoon noch endergoon. Evenwicht situatie

16
De Gibbs vrije energie, DG
DG = Gproducten – G reactanten DG is onafhankelijk van de gekozen route DG zegt niets over de snelheid van de reactie Indien negatief: dan kan de reactie plaats vinden. Dat wil niet zeggen dat het merkbaar of snel gebeurt!

17
Relatie tussen vrije energie en evenwichtsconstante K
DG = DG0 + RT ln K K is quotiënt van de concentraties reactanten K = [AB]/[A][B] R is de gasconstante DG0 is de standaard vrije energie (vaste waarde)
")
18
Allosterische enzymen: sigmoidale curve

19
Allosterische enzymen
Binden een bepaald molecule Veranderen dan van vorm Kunnen nu efficiënt aan substraat binden Hemoglobine kent een allosterisch effect door zuurstof binding

20
Gekoppelde enzymatische reacties

21
Co-factoren, co-enzymen en prostetische groep
Veel enzymen hebben andere verbindingen nodig Zijn klein t.o.v. het eiwit Anorganisch: Co-factor Organisch: Covalent: prostetische groep Niet-covalent: co-enzym (vaak afgeleid van vitamines)
")
22
Prosthetische groepen en cofactoren: zijn nodig voor enzymatische activiteit

23
Katalytisch vermogen Enzymen kunnen reacties versnellen tot x zo snel als niet-gekatalyseerde reacties! Urease is een goed voorbeeld: gekatalyseerd: 3x104/sec Niet gekatalyseerd: 3x10-10/sec Ratio is 1x1014 !

24
Specificiteit Enzymen herkennen hun substraat (substraat-specificiteit) Enzymen leveren een opbrengst van meer dan 95% (reactiespecificiteit) Specificiteit wordt bepaald door de unieke “fit” tussen het substraat en het enzym

25
Enzymatische modellen

26
De cyclus van een enzym

27
Enzymatische werking

28
Schematisch de werking van een enzym

29
Een voorbeeld van een enzym

30
Wat enzymen doen.... Enzymen versnellen reacties door verlaging van de activeringsenergie Enzymen doen dit door de “transition state” van de reactie beter te binden dan het substraat

31
enzymreacties Foutje in tekstboek: DP is niet negatief, want wordt gevormd!

32
Enzymreacties k1 k2 E + S ES E + P k-1
E is enzym S is substraat ES is het complex van E en S P is product k zijn snelheidsconstantes K is (k-1 + k2) / k1
 / k1.")
33
Verzadiging Bij kleine hoeveelheid enzym t.o.v. substraat
Enzym op maximale snelheid Enzym kan niet sneller omzetten Verzadiging van het enzym

34
Reactiesnelheid is afhankelijk van de concentratie substraat

35
Bij hoge S concentraties nadert v de Vmax waarde

36
Michaelis - Menten Leonor Michaelis Maud Menten

37
Michaelis Menten kinetiek
De basis voor de meeste niet-allosterische enzym-reacties Ontwikkeld in 1913 Karakteriseert enzym-activiteit in termen van snelheid en binding aan het substraat

38
Reactieschema k1 k2 E + S ES E + P k-1

39
Michaelis-Menten vergelijking
Michaelis-Menten's theorie Gaat uit van de vorming van een enzymsubstraatcomplex ES is in een snel evenwicht met E en S De reactie van ES naar E en P gaat langzaam

40
Eerste orde kinetiek: er is een lineair verband tussen enzymatische snelheid en substraat concentratie

41
Michaelis-Menten mechanisme
snelheid vorming ES = k1.[E].[S] snelheid van afbraak van ES = (k-1+k2).[ES] a Km = [E].[S] = (k-1+k2)/k1 [ES] Km = Michaelis-Menten constante
.[ES] a Km = [E].[S] = (k-1+k2)/k1. [ES] Km = Michaelis-Menten constante.")
42
Michaelis-Menten – vergelijking: Vmax
Michaelis-Menten – vergelijking: Vmax.[S] v = Km + [S]

43
De Vmax Vmax is een “constante”
Vmax is de theoretisch maximale snelheid van de reactie Vmax vereist dat alle enzymmoleculen aan het substraat gebonden zijn Vmax wordt benaderd als de substraatconcentratie hoog is

44
De steady state Er is weinig enzym-substraat complex aanwezig
Complex vorming en afbraak zijn in evenwicht Enzymen bereiken snel de steady state fase: zijn efficiënt in het katalyseren van de reactie

45
Een maat voor de katalytische activiteit
Het turnover getal Een maat voor de katalytische activiteit is het aantal substraatmoleculen omgezet in product per enzymmolecuul per sec, als het enzym (E) is verzadigd met substraat. k2 = kcat = Vmax/[Et] kcat varieert per enzym (minder dan 1/sec tot vele miljoenen/sec
 is verzadigd met substraat. k2 = kcat = Vmax/[Et] kcat varieert per enzym (minder dan 1/sec tot vele miljoenen/sec.")
46
Enzymen beschrijven we middels kcat en Km

47
De Michaelisconstante Km
Km is een constante waarbij de Vmax half-maximaal is Km is afgeleid van de reactiesnelheid-constanten Kleine Km: sterke binding; hoge Km: zwakke binding

48
Bepaling Km

49
Lineaire Plot van Michaelis-Mentenvergelijking
V0 = Vmax.[S]/(KM + [S]) Lineweaver-Burk: zet in reciproke vorm 1/V0 = (KM/Vmax).1/[S] + 1/Vmax Vergelijk met : y = a.x + b
 Lineweaver-Burk: zet in reciproke vorm. 1/V0 = (KM/Vmax).1/[S] + 1/Vmax. Vergelijk met : y = a.x + b.")
50
Door de Michaelis Menten curve lineair te maken zijn Km en Vmax eenvoudig te bepalen
Lineweaver-Burk-plot

52
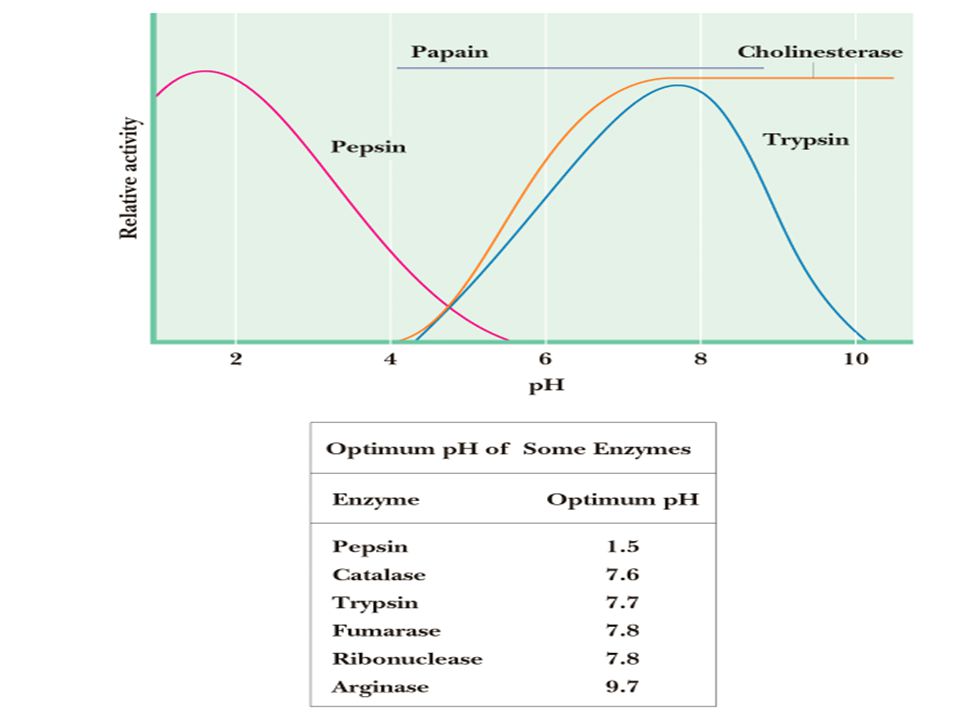
Invloed pH

53
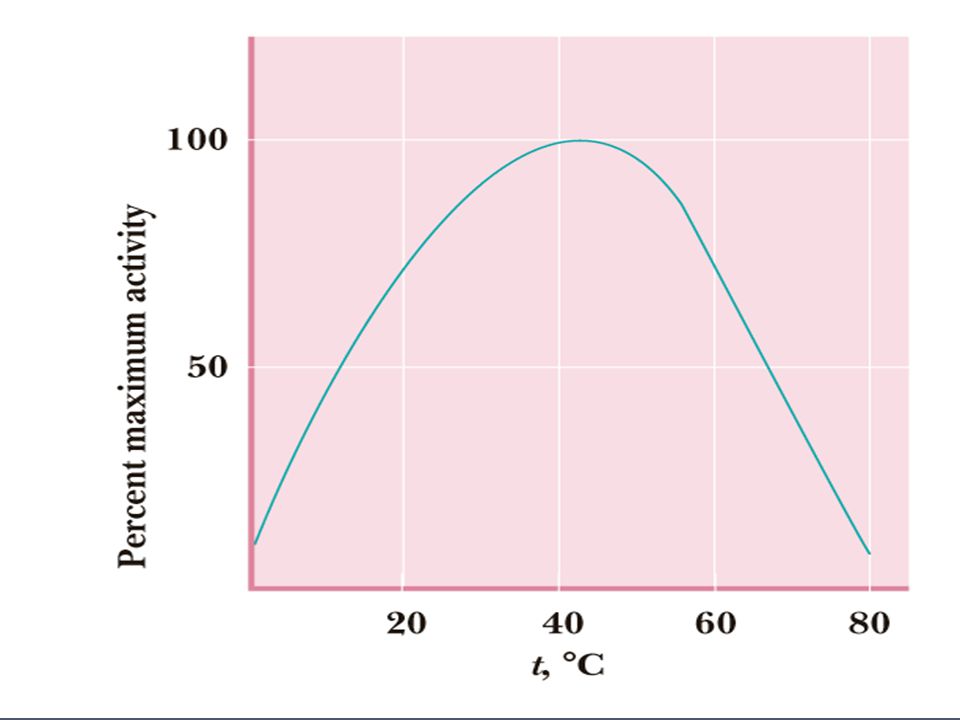
Invloed temperatuur Hogere temperatuur:
Moleculen bewegen sneller Reactie verloopt sneller Te hoge temperatuur: enzym denatureert Dus enzymen hebben een optimum

55
Invloed temperatuur b.v. Taq DNA polymerase

56
Enzym inhibitie Enzymen kunnen specifiek geremd worden
Farma bedrijven proberen soms een miljoen “lead compounds” Uiteindelijk blijft er 1 over die precies past in het enzym en specifiek de activiteit remt Ontwikkeling kost soms miljoenen euros Terugverdienen via octrooirecht op het uiteindelijke geneesmiddel Ethisch: derde wereld ziekten en ontwikkeling medicijnen Medicijnen zijn soms heel duur

57
Reversibel versus Irreversibel
Enzymremmers Reversibel versus Irreversibel Reversibele inhibitors: niet-covalente binding Irreversibele inhibitors: (bijna) covalente binding
 covalente binding.")
58
Remming van enzymen Competitief - inhibitor (I) bindt aan actieve plaats Niet-competitief- inhibitor (I) bindt elders
 bindt elders.")
59
Competitieve remming Een remmer bindt op de plaats van het substraat
Er is meer substraat nodig om de remming op te heffen De Km wordt dus groter

60
Competitieve remming Competitieve remming

61
Niet-competitieve remming
Het enzym wordt ten dele geremd doordat een remmer op een andere plaats bindt dan het substraat, maar wel de katalyse beinvloed

62
Niet-competitieve remming

63
De Michaelis curve bij remming

64
Irreversibele remming
Remmer bindt zeer sterk aan het enzym en laat vrijwel niet meer los Zowel remmer als enzym gaan dus verloren in het irreversibele complex

65
Enzymactiviteit bepalen...Hoe?
Kies de de substraatconcentratie......? het substraat.....? de pH ? de aard van de buffer.....? de temperatuur.....?

66
Enzymactiviteit bepalen...Waarom?
Diagnostiek: veel eiwitten spelen een rol in medische situaties Eiwitzuiveringen, isolaties, research Met hoeveel enzym(in Units) begin je? Hoeveel enzym (in Units) heb je nog over? Hoe zuiver (U/mg) is het?
 begin je Hoeveel enzym (in Units) heb je nog over Hoe zuiver (U/mg) is het")
67
Diagnostiek Enzym Normale Waarden (IU/l) Verhoogd wijst op
Creatine- fosfokinase (man) (vrouw) Hartinfarct Alkalische fosfatase 30-125 galstuwing Zure fosfatase 2,5 Prostaat- carcinoom Lactaatdehydro- genase leverziekten
 (vrouw) Hartinfarct. Alkalische fosfatase galstuwing. Zure fosfatase. 2,5. Prostaat- carcinoom. Lactaatdehydro- genase leverziekten.")
68
Waarom Units en geen mol/l?
Eiwitten ingewikkeld mengsel Eiwitten kunnen denatureren Kijk daarom naar de activiteit van het enzym!!! 1 I.U = die hoev.enzym die in staat is om 1 mol substraat/min om te zetten (onder bepaalde condities)
")
69
Voorbeeld enzymbepaling
Zorg voor: Substraat met kleine KM, goede affiniteit Hoge [S] Optimale temperatuur Optimale pH Optimale buffer En meet A/min en activiteit enzym

70
Km en Kcat Belangrijkste parameters voor enzymen
Hiermee karakteriseren we enzymen! Km: affiniteit voor substraat en = ½ Vmax Kcat: turnover getal, dus mate van katalytisch vermogen ten opzichte van het substraat

71
Voorbeeld: enzym remming bij AIDS
Tegen drie cruciale virale enzymen zijn remmers ontworpen De enzymen zijn “gekristalliseerd”, dwz, de structuur is onderzocht en bekend Er zijn “gericht” medicijnen gemaakt tegen deze enzymen

72
Virale enzymen en AIDS

73
Anti-virale middelen en enzymatische targeting

74
Samenvatting Enzymen zijn zeer efficiënte katalytische eiwitten
Michaelis en Menten beschreven een zeer goed model voor niet-allosterische enzym activiteit Uit een Michaelis Menten curve kan men de Km bepalen Kcat kan berekend worden uit de hoeveelheid gevormd product (is k2) Enzymen kunnen competitief niet-competitief geremd worden Bij competitieve remming verandert de Km Bij niet-competitieve remming verandert de Vmax
 Enzymen kunnen competitief niet-competitief geremd worden. Bij competitieve remming verandert de Km. Bij niet-competitieve remming verandert de Vmax.")
Verwante presentaties
>")


















