Download de presentatie
1
Gemeenschappen

2
Gemeenschapsstructuur

3
Gemeenschapsstructuur
Onder natuurlijke omstandigheden maken soorten deel uit van complexe gemeenschappen, d.w.z. populaties van verschillende soorten die met elkaar interageren tijdens dezelfde periode en op dezelfde plaats.

4
Gemeenschapsstructuur
Onder natuurlijke omstandigheden maken soorten deel uit van complexe gemeenschappen, d.w.z. populaties van verschillende soorten die met elkaar interageren tijdens dezelfde periode en op dezelfde plaats. Gemeenschappen vertonen kenmerken die we niet aan-treffen bij de afzonderlijke soorten waaruit ze zijn samen-gesteld.

5
Gemeenschapsstructuur
Onder natuurlijke omstandigheden maken soorten deel uit van complexe gemeenschappen, d.w.z. populaties van verschillende soorten die met elkaar interageren tijdens dezelfde periode en op dezelfde plaats. Gemeenschappen vertonen kenmerken die we niet aan-treffen bij de afzonderlijke soorten waaruit ze zijn samen-gesteld. Hun structuur en functioneren wordt bepaald door abio-tische en biotische factoren.

6
Patronen van diversiteit

7
Patronen van diversiteit

8
Patronen van diversiteit

9
Patronen van diversiteit
Gemeenschapsecologen hanteren verschillende concepten om diversiteit te beschrijven en kwantificeren:

10
Patronen van diversiteit
Gemeenschapsecologen hanteren verschillende concepten om diversiteit te beschrijven en kwantificeren: α-diversiteit: diversiteit binnen een welbepaalde gemeen-schap, uitgedrukt als aantal soorten.

11
Patronen van diversiteit
Gemeenschapsecologen hanteren verschillende concepten om diversiteit te beschrijven en kwantificeren: α-diversiteit: diversiteit binnen een welbepaalde gemeen-schap, uitgedrukt als aantal soorten. β-diversiteit: verandering in diversiteit tussen gemeen-schappen, uitgedrukt als aantal unieke soorten.

12
Patronen van diversiteit
Gemeenschapsecologen hanteren verschillende concepten om diversiteit te beschrijven en kwantificeren: α-diversiteit: diversiteit binnen een welbepaalde gemeen-schap, uitgedrukt als aantal soorten. β-diversiteit: verandering in diversiteit tussen gemeen-schappen, uitgedrukt als aantal unieke soorten. γ-diversiteit: diversiteit over alle gemeenschappen in een regio, uitgedrukt als totaal aantal soorten.

13
Patronen van diversiteit
Strikt genomen houdt de term diversiteit tevens rekening met het relatieve ‘belang’ van elke soort in een gemeen-schap uitgedrukt als abundantie, productiviteit, grootte, etc.

14
Patronen van diversiteit
Strikt genomen houdt de term diversiteit tevens rekening met het relatieve ‘belang’ van elke soort in een gemeen-schap uitgedrukt als abundantie, productiviteit, grootte, etc. Mathematisch vertaald omvat diversiteit een combinatie van soortenrijkdom en eveness, het relatieve aandeel van elke soort in een gemeenschap.

15
Patronen van diversiteit
Strikt genomen houdt de term diversiteit tevens rekening met het relatieve ‘belang’ van elke soort in een gemeen-schap uitgedrukt als abundantie, productiviteit, grootte, etc. Mathematisch vertaald omvat diversiteit een combinatie van soortenrijkdom en eveness, het relatieve aandeel van elke soort in een gemeenschap.

16
Patronen van diversiteit

17
Eilandbiogeografie

18
Eilandbiogeografie The ‘Equilibrium Theory of Island Biogeography’ (Robert MacArthur & Edward Wilson, 1967) .
 .")
19
Eilandbiogeografie The ‘Equilibrium Theory of Island Biogeography’ (Robert MacArthur & Edward Wilson, 1967) . Gemiddeld lagere soortenrijkdom op eilanden t.o.v. ver-gelijkbare gebieden op het vasteland.

20
Eilandbiogeografie vogels Azoren planten Azoren species-area verbanden
reptielen West-Indies vogels Solomon

21
Eilandbiogeografie zoogdierenbergketens broedvogels bosfragmenten
species-area verbanden invertebraten mosselbanken vissen woestijnbronnen

22
Eilandbiogeografie

23
Eilandbiogeografie The ‘Equilibrium Theory of Island Biogeography’ (Robert MacArthur & Edward Wilson, 1967) . Gemiddeld lagere soortenrijkdom op eilanden t.o.v. ver-gelijkbare gebieden op het vasteland. Soortenrijkdom op oceanische eilanden vertoont een dyna-misch evenwicht tussen ratios van immigratie en emigratie.

24
Eilandbiogeografie

25
Eilandbiogeografie

26
Eilandbiogeografie

27
Functioneren van gemeenschappen

28
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten).
 of sleutelrol (keystone soorten).")
29
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Dominante soorten
 of sleutelrol (keystone soorten). Dominante soorten.")
30
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Dominante soorten hoge abundantie of biomassa
 of sleutelrol (keystone soorten). Dominante soorten. hoge abundantie of biomassa.")
31
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Dominante soorten hoge abundantie of biomassa hoge impact op voorkomen of verspreiding van andere soorten en op abiotische variabelen
 of sleutelrol (keystone soorten). Dominante soorten. hoge abundantie of biomassa. hoge impact op voorkomen of verspreiding van andere. soorten en op abiotische variabelen.")
32
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Dominante soorten hoge abundantie of biomassa hoge impact op voorkomen of verspreiding van andere soorten en op abiotische variabelen hoog competitief vermogen
 of sleutelrol (keystone soorten). Dominante soorten. hoge abundantie of biomassa. hoge impact op voorkomen of verspreiding van andere. soorten en op abiotische variabelen. hoog competitief vermogen.")
33
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Keystone soorten
 of sleutelrol (keystone soorten). Keystone soorten.")
34
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Keystone soorten niet noodzakelijk hoge abundantie of biomassa
 of sleutelrol (keystone soorten). Keystone soorten. niet noodzakelijk hoge abundantie of biomassa.")
35
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Keystone soorten niet noodzakelijk hoge abundantie of biomassa hoge impact via ecologische rol of niche
 of sleutelrol (keystone soorten). Keystone soorten. niet noodzakelijk hoge abundantie of biomassa. hoge impact via ecologische rol of niche.")
36
Functioneren van gemeenschappen
Sommige soorten hebben een grote invloed op structuur of functioneren van gemeenschappen wegens hun aantal (dominante soorten) of sleutelrol (keystone soorten). Keystone soorten niet noodzakelijk hoge abundantie of biomassa hoge impact via ecologische rol of niche belang blijkt vooral na het verdwijnen
 of sleutelrol (keystone soorten). Keystone soorten. niet noodzakelijk hoge abundantie of biomassa. hoge impact via ecologische rol of niche. belang blijkt vooral na het verdwijnen.")
37
Functioneren van gemeenschappen

38
Functioneren van gemeenschappen

39
Voedselketens en voedselwebben

40
Voedselketens en voedselwebben
Voedselrelaties vormen een zeer belangrijke component in de structuur en dynamiek van gemeenschappen.

41
Voedselketens en voedselwebben
Voedselrelaties vormen een zeer belangrijke component in de structuur en dynamiek van gemeenschappen. Het complexe geheel van voedselrelaties wordt trofische gemeenschapsstructuur genoemd.

42
Voedselketens en voedselwebben
Voedselrelaties vormen een zeer belangrijke component in de structuur en dynamiek van gemeenschappen. Het complexe geheel van voedselrelaties wordt trofische gemeenschapsstructuur genoemd. Primaire producenten (autotrofen) capteren zonlicht en vormen via fotosynthese complexe organische moleculen uit H2O en CO2.
 capteren zonlicht en vormen via fotosynthese complexe organische moleculen uit H2O en CO2.")
43
Voedselketens en voedselwebben
Voedselrelaties vormen een zeer belangrijke component in de structuur en dynamiek van gemeenschappen. Het complexe geheel van voedselrelaties wordt trofische gemeenschapsstructuur genoemd. Primaire producenten (autotrofen) capteren zonlicht en vormen via fotosynthese complexe organische moleculen uit H2O en CO2. Primaire consumenten (heterotrofen) prederen op auto-trofen, secundaire heterotrofen op primaire heterotrofen, etc. Naargelang het dieet onderscheiden we herbivoren, carnivoren, omnivoren, detrivoren en saprofieten.
 capteren zonlicht en vormen via fotosynthese complexe organische moleculen uit H2O en CO2. Primaire consumenten (heterotrofen) prederen op auto-trofen, secundaire heterotrofen op primaire heterotrofen, etc. Naargelang het dieet onderscheiden we herbivoren, carnivoren, omnivoren, detrivoren en saprofieten.")
44
Voedselketens en voedselwebben
Voedselketen: Lineaire energiestroom waarbij energie als voedsel meerdere trofische niveaus doorloopt.

45
Voedselketens en voedselwebben

46
Voedselketens en voedselwebben
Voedselketen: Lineaire energiestroom waarbij energie als voedsel meerdere trofische niveaus doorloopt. Voedselweb: Naast directe interacties tussen trofische niveaus worden trofische gemeenschappen ook geken-merkt door indirecte interacties binnen trofische niveaus.

47
Voedselketens en voedselwebben

48
Voedselketens en voedselwebben
Voedselketen: Lineaire energiestroom waarbij energie als voedsel meerdere trofische niveaus doorloopt. Voedselweb: Naast directe interacties tussen trofische niveaus worden trofische gemeenschappen ook geken-merkt door indirecte interacties binnen trofische niveaus. Bij elke overgang tussen trofische niveaus gaat ca. 90% van de energie opgeslagen in organische componenten verloren.

49
Voedselketens en voedselwebben

50
Voedselketens en voedselwebben
Het aantal trofische niveaus in voedselketens is beperkt.

51
Voedselketens en voedselwebben
Het aantal trofische niveaus in voedselketens is beperkt. energetische hypothese

52
Voedselketens en voedselwebben
Het aantal trofische niveaus in voedselketens is beperkt. energetische hypothese dynamische stabiliteit hypothese

53
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld:

54
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld: Bottom-up model

55
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld: Bottom-up model verandering in biomassa van lagere tropische niveaus indiceert veranderingen op hogere niveaus

56
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld: Bottom-up model verandering in biomassa van lagere trofische niveaus indiceert veranderingen op hogere niveaus weergegeven als N V H P

57
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld: Top-down model (trofisch cascade model)
")
58
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld: Top-down model (trofisch cascade model) structuur van een gemeenschap wordt in hoofdzaak ge-reguleerd door predatie
 structuur van een gemeenschap wordt in hoofdzaak ge-reguleerd door predatie.")
59
Gemeenschapsregulatie
Op basis van potentiële trofische interacties worden twee modellen van gemeenschapsregulatie voorgesteld: Top-down model (trofisch cascade model) structuur van een gemeenschap wordt in hoofdzaak ge-reguleerd door predatie weergegeven als N V H P
 structuur van een gemeenschap wordt in hoofdzaak ge-reguleerd door predatie. weergegeven als N V H P.")
60
Gemeenschapsregulatie
Traditioneel bestond de visie dat gemeenschappen zich in afwezigheid van verstoring in evenwicht bevinden, d.w.z. een tendens vertonen tot constante soortensamenstelling.

61
Gemeenschapsregulatie
Traditioneel bestond de visie dat gemeenschappen zich in afwezigheid van verstoring in evenwicht bevinden, d.w.z. een tendens vertonen tot constante soortensamenstelling. Momenteel primeert de visie van een niet-evenwichts-model (non-equilibrium model).
.")
62
Gemeenschapsregulatie
Traditioneel bestond de visie dat gemeenschappen zich in afwezigheid van verstoring in evenwicht bevinden, d.w.z. een tendens vertonen tot constante soortensamenstelling. Momenteel primeert de visie van een niet-evenwichts-model (non-equilibrium model). Sommige gemeenschappen zijn voor hun voortbestaan (gedeeltelijk) afhankelijk van verstoringen.
. Sommige gemeenschappen zijn voor hun voortbestaan (gedeeltelijk) afhankelijk van verstoringen.")
64
Gemeenschapsregulatie
Traditioneel bestond de visie dat gemeenschappen zich in afwezigheid van verstoring in evenwicht bevinden, d.w.z. een tendens vertonen tot constante soortensamenstelling. Momenteel primeert de visie van een niet-evenwichts-model (non-equilibrium model). Sommige gemeenschappen zijn voor hun voortbestaan (gedeeltelijk) afhankelijk van verstoringen. Intermediate disturbance hypothesis: middelmatige verstoring leidt tot de hoogste soortenrijkdom (Joseph Connell, 1978).
. Sommige gemeenschappen zijn voor hun voortbestaan (gedeeltelijk) afhankelijk van verstoringen. Intermediate disturbance hypothesis: middelmatige verstoring leidt tot de hoogste soortenrijkdom (Joseph Connell, 1978).")
65
Gemeenschapsregulatie
Joseph Connell Research Professor University of California, Santa Barbara

66
Gemeenschapsregulatie
Na grootschalige verstoring zullen nieuwe gemeenschap-pen zich geleidelijk en via opeenvolgende stadia (elk gekarakteriseerd door dominante soorten) ontwikkelen (ecologische successie).
 ontwikkelen (ecologische successie).")
67
Gemeenschapsregulatie
Na grootschalige verstoring zullen nieuwe gemeenschap-pen zich geleidelijk en via opeenvolgende stadia (elk gekarakteriseerd door dominante soorten) ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap.
 ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap.")
68
Gemeenschapsregulatie
Na grootschalige verstoring zullen nieuwe gemeenschap-pen zich geleidelijk en via opeenvolgende stadia (elk gekarakteriseerd door dominante soorten) ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen:
 ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen:")
69
Gemeenschapsregulatie
Na grootschalige verstoring zullen nieuwe gemeenschap-pen zich geleidelijk en via opeenvolgende stadia (elk gekarakteriseerd door dominante soorten) ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen: Facilitatie: vroege soorten bespoedigen vestiging
 ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen: Facilitatie: vroege soorten bespoedigen vestiging.")
70
Gemeenschapsregulatie
Na grootschalige verstoring zullen nieuwe gemeenschap-pen zich geleidelijk en via opeenvolgende stadia (elk gekarakteriseerd door dominante soorten) ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen: Facilitatie: vroege soorten bespoedigen vestiging Inhibering: vroege soorten bemoeilijken vestiging
 ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen: Facilitatie: vroege soorten bespoedigen vestiging. Inhibering: vroege soorten bemoeilijken vestiging.")
71
Gemeenschapsregulatie
Na grootschalige verstoring zullen nieuwe gemeenschap-pen zich geleidelijk en via opeenvolgende stadia (elk gekarakteriseerd door dominante soorten) ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen: Facilitatie: vroege soorten bespoedigen vestiging Inhibering: vroege soorten bemoeilijken vestiging Tolerantie: vroege soorten hebben een neutrale invloed
 ontwikkelen (ecologische successie). Speelt zich af op ecologische (niet evolutionaire) tijd-schalen en leidt tot dynamische climax gemeenschap. Successie wordt gestuurd door drie ecologische processen: Facilitatie: vroege soorten bespoedigen vestiging. Inhibering: vroege soorten bemoeilijken vestiging. Tolerantie: vroege soorten hebben een neutrale invloed.")
72
Gemeenschapsregulatie
Primaire successie

73
Gemeenschapsregulatie
Primaire successie temporele verandering in soortensamenstelling in voor- heen onbewoond gebied

74
Gemeenschapsregulatie

75
Gemeenschapsregulatie
Primaire successie temporele verandering in soortensamenstelling in voor- heen onbewoond gebied windverbreide varens en mossen vormen de eerste fotosynthetische kolonisten

76
Gemeenschapsregulatie
Primaire successie temporele verandering in soortensamenstelling in voor- heen onbewoond gebied windverbreide varens en mossen vormen de eerste fotosynthetische kolonisten decompositie leidt tot de vorming van een initiële organische bodemlaag

77
Gemeenschapsregulatie
Secundaire successie

78
Gemeenschapsregulatie
Secundaire successie temporele verandering in soortensamenstelling nadat originele vegetatie verdween (maar intacte bodem)
")
79
Gemeenschapsregulatie

80
Gemeenschapsregulatie
Secundaire successie temporele verandering in soortensamenstelling nadat originele vegetatie verdween (maar intacte bodem) via successie ontstaat een vegetatie die sterk gelijkt op de oorspronkelijke
 via successie ontstaat een vegetatie die sterk gelijkt op de oorspronkelijke.")
81
Gemeenschapsregulatie

82
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen.

83
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Interactief gemeenschapsmodel

84
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Interactief gemeenschapsmodel gemeenschappen gedragen zich als ‘superorganisme’

85
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Interactief gemeenschapsmodel gemeenschappen gedragen zich als ‘superorganisme’ vertonen ‘ontwikkeling’ tot ‘volwassen’ staat

86
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Interactief gemeenschapsmodel gemeenschappen gedragen zich als ‘superorganisme’ vertonen ‘ontwikkeling’ tot ‘volwassen’ staat nadruk op interactie tussen soorten

87
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Individualistisch gemeenschapsmodel

88
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Individualistisch gemeenschapsmodel omgevingsgradiënten belangrijker dan interacties

89
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Individualistisch gemeenschapsmodel omgevingsgradiënten belangrijker dan interacties samen voorkomen duidt op gelijkaardige vereisten

90
Gemeenschapsmodellen
Vraag of gemeenschappen op een ‘voorspelbare’ manier gestructureerd zijn, blijft bron van discussie onder ge-meenschapsecologen. Individualistisch gemeenschapsmodel omgevingsgradiënten belangrijker dan interacties samen voorkomen duidt op gelijkaardige vereisten nadruk op individualiteit van soorten

91
Gemeenschapsmodellen
Interactief

92
Gemeenschapsmodellen
Interactief Structuur en dynamiek Individua-listisch

93
Ecologische nicheconcept

94
Ecologische nicheconcept
Ecologische niche omvat alle biotische en abiotische aspecten die deel uitmaken van de levensbehoefte van een organisme.

95
Ecologische nicheconcept

96
Ecologische nicheconcept
Ecologische niche omvat alle biotische en abiotische aspecten die deel uitmaken van de levensbehoefte van een organisme. In evolutionaire termen weerspiegelt het de totaliteit aan adaptaties van een organisme aan zijn leefomgeving.

97
Ecologische nicheconcept
Ecologische niche omvat alle biotische en abiotische aspecten die deel uitmaken van de levensbehoefte van een organisme. In evolutionaire termen weerspiegelt het de totaliteit aan adaptaties van een organisme aan zijn leefomgeving. Limiterende hulpbron (critical resource): elke omgevings-component die de ecologische niche van een organisme beperkt.
: elke omgevings-component die de ecologische niche van een organisme beperkt.")
98
Ecologische nicheconcept
Ecologische niche omvat alle biotische en abiotische aspecten die deel uitmaken van de levensbehoefte van een organisme. In evolutionaire termen weerspiegelt het de totaliteit aan adaptaties van een organisme aan zijn leefomgeving. Limiterende hulpbron (critical resource): elke omgevings-component die de ecologische niche van een organisme beperkt. Gerealiseerde niche is het deel van de fundamentele (potentiële) niche dat effectief wordt ingenomen gegeven competitie voor limiterende hulpbronnen.
: elke omgevings-component die de ecologische niche van een organisme beperkt. Gerealiseerde niche is het deel van de fundamentele (potentiële) niche dat effectief wordt ingenomen gegeven competitie voor limiterende hulpbronnen.")
99
Ecologische nicheconcept
Chthamalus Balanus

100
Ecologische nicheconcept

101
Ecologische nicheconcept
Het nicheconcept is nauw verweven met het verschijnsel competitie (interactie die optreedt wanneer twee of meer individuen gebruik willen maken van dezelfde kritische hulpbron).
.")
102
Ecologische nicheconcept
Het nicheconcept is nauw verweven met het verschijnsel competitie (interactie die optreedt wanneer twee of meer individuen gebruik willen maken van dezelfde kritische hulpbron). contest competition: impliceert actieve interferentie
. contest competition: impliceert actieve interferentie.")
103
Ecologische nicheconcept
Het nicheconcept is nauw verweven met het verschijnsel competitie (interactie die optreedt wanneer twee of meer individuen gebruik willen maken van dezelfde kritische hulpbron). contest competition: impliceert actieve interferentie scramble competition: exploitatie van eenzelfde hulpbron
. contest competition: impliceert actieve interferentie. scramble competition: exploitatie van eenzelfde hulpbron.")
104
Ecologische nicheconcept
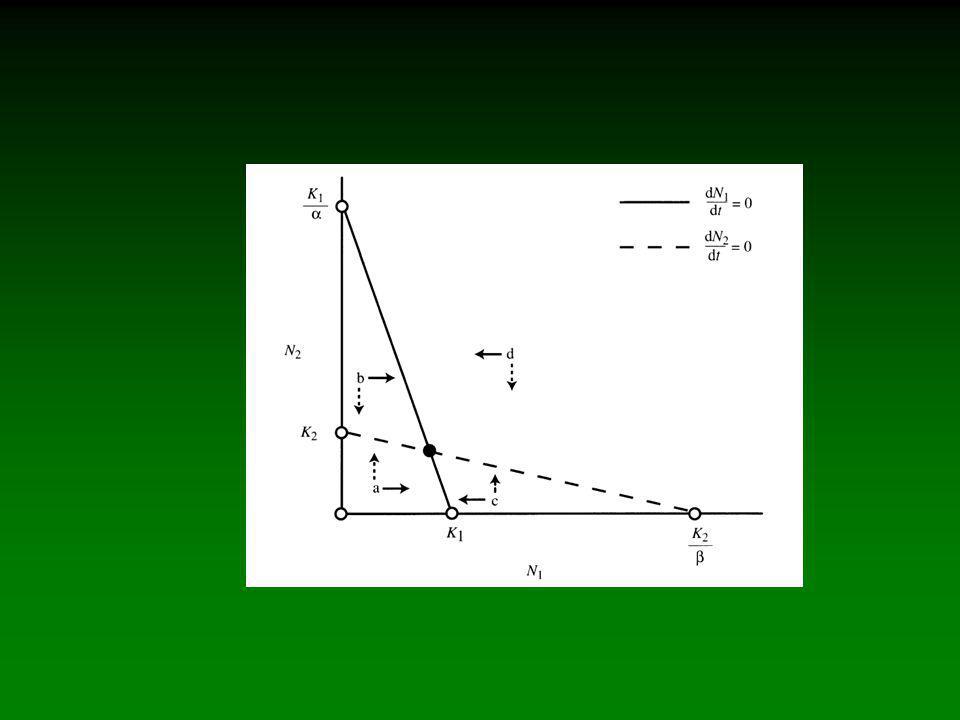
Het nicheconcept is nauw verweven met het verschijnsel competitie (interactie die optreedt wanneer twee of meer individuen gebruik willen maken van dezelfde kritische hulpbron). contest competition: impliceert actieve interferentie scramble competition: exploitatie van eenzelfde hulpbron Competitie voor limiterende hulpbronnen kan aanleiding geven tot competitieve uitsluiting (G.F. Gause, 1934).
. contest competition: impliceert actieve interferentie. scramble competition: exploitatie van eenzelfde hulpbron. Competitie voor limiterende hulpbronnen kan aanleiding geven tot competitieve uitsluiting (G.F. Gause, 1934).")
105
Ecologische nicheconcept
G.F. Gause Moscow University

108
Ecologische nicheconcept
Twee soorten van eenzelfde gemeenschap kunnen niet exact dezelfde ecologische niche bezetten.

109
Ecologische nicheconcept
Twee soorten van eenzelfde gemeenschap kunnen niet exact dezelfde ecologische niche bezetten. Over een evolutionaire tijdschaal zal natuurlijke selectie individuen bevoordelen die competitie voor kritische hulp-bonnen reduceren.

112
Niche dimensie Gebruik hulpbron

113
Niche dimensie Gebruik hulpbron Niche dimensie Gebruik hulpbron

114
Ecologische nicheconcept
Twee soorten van eenzelfde gemeenschap kunnen niet exact dezelfde ecologische niche bezetten. Over een evolutionaire tijdschaal zal natuurlijke selectie individuen bevoordelen die competitie voor kritische hulp-bonnen reduceren. Resource partitioning: reductie in competitie tussen co-existerende soorten door reductie van overlap in één of meerdere niche dimensies. Dit proces kan leiden tot character displacement.

115
Ecologische nicheconcept

116
Andere biotische interacties

117
Andere biotische interacties
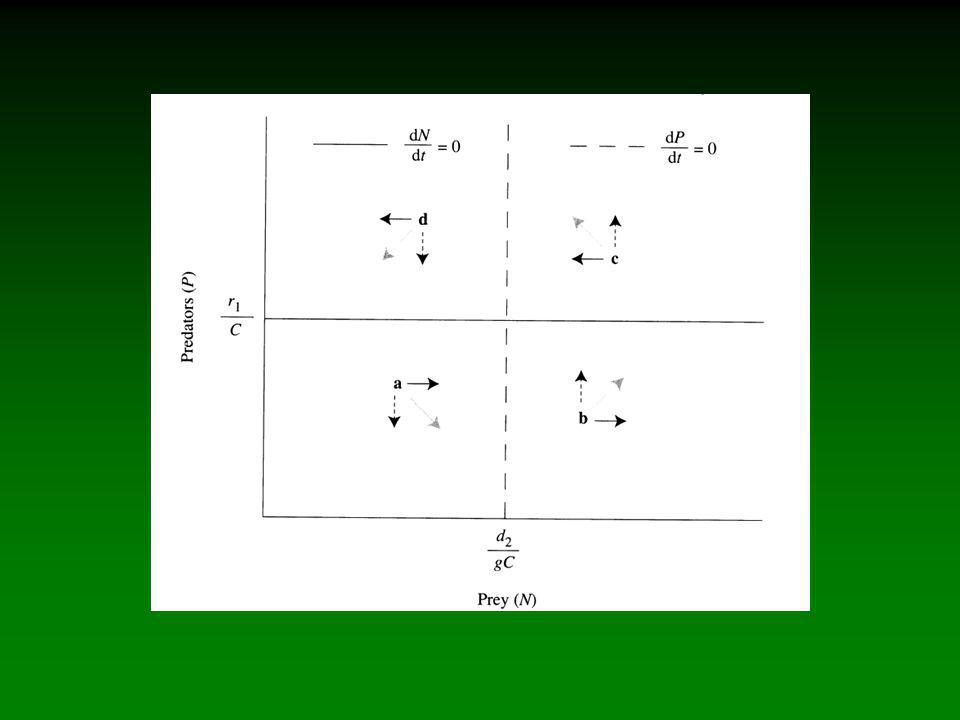
Predatie verwijst naar het proces waarbij een prooi geconsumeerd wordt door een predator. Dit slaat zowel op carnivoren áls herbivoren.

120
Andere biotische interacties
Predatie verwijst naar het proces waarbij een prooi geconsumeerd wordt door een predator. Dit slaat zowel op carnivoren áls herbivoren. Natuurlijke selectie leidt tot verfijning van adaptaties bij predatoren en prooien. Evolutie van strategieën bij de ene groep vormt de selectieve kracht voor co-evolutie van strategieën bij de tegenpartij (evolutionaire wapenwedloop).
.")
121
Andere biotische interacties
Predatie verwijst naar het proces waarbij een prooi geconsumeerd wordt door een predator. Dit slaat zowel op carnivoren áls herbivoren. Natuurlijke selectie leidt tot verfijning van adaptaties bij predatoren en prooien. Evolutie van strategieën bij de ene groep vormt de selectieve kracht voor co-evolutie van strategieën bij de tegenpartij (evolutionaire wapenwedloop). Adaptaties kunnen zowel morfologisch (bv. kleurpatronen), fysiologisch (bv. alkaloïden) als gedragsmatig (bv. groeps-gedrag) zijn.
. Adaptaties kunnen zowel morfologisch (bv. kleurpatronen), fysiologisch (bv. alkaloïden) als gedragsmatig (bv. groeps-gedrag) zijn.")
122
aposematisme

123
Mülleriaanse mimicry

124
Batesiaanse mimicry

125
Andere biotische interacties
Symbiose verwijst naar een nauwe associatie tussen indi-viduen van twee of meer soorten als gevolg van co-evolutie. Afhankelijk van het type van symbiose kunnen symbionten voordeel, nadeel of neutraliteit ondervinden.

126
Andere biotische interacties
Effect op 1 Effect op 2 Predatie van 2 door 1 positief negatief Mutualisme van 1 en 2 neutraal Parasitisme van 1 op 2 Competitie van 1 en 2 Commensalisme van 1 & 2

127
Mutualisme

128
Andere biotische interacties
Effect op 1 Effect op 2 Predatie van 2 door 1 positief negatief Mutualisme van 1 en 2 neutraal Parasitisme van 1 op 2 Competitie van 1 en 2 Commensalisme van 1 & 2

129
Commensalisme

130
Andere biotische interacties
Effect op 1 Effect op 2 Predatie van 2 door 1 positief negatief Mutualisme van 1 en 2 neutraal Parasitisme van 1 op 2 Competitie van 1 en 2 Commensalisme van 1 & 2

131
Parasitisme