Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Expressievectoren en proteïneproductiesystemen
o.a. Primrose & Twyman, 7e editie, pp.81-93 Primrose, Twyman & Old, 6e editie, pp.70-83 Expressievectoren en proteïneproductiesystemen Expressie verwijst het gen, productie verwijst naar het proteïne, hoewel in het jargon 'expressie' ook vaak verkeerdelijk voor proteïnen wordt misbruikt. Aangezien productie van een gekloneerd proteïne in hoge hoeveelheden gebeurt, is detectie en analyse van het rendement meestal relatief eenvoudig, en kan dit makkelijk door visualisering (kleuring) op SDS-PAGE. Hierbij kan expressie "voor" en "na" inductie vergeleken worden, evenals het celcompartiment of de toestand waarin het eiwitproduct zich bevindt : cytoplasmatisch versus periplasmatisch versus extracellulair, en oplosbaar of onoplosbaar. (Dit laatste aspect is niet in het voorbeeld van volgende figuur opgenomen.)
 op SDS-PAGE. Hierbij kan expressie voor en na inductie vergeleken worden, evenals het. celcompartiment of de toestand waarin het eiwitproduct zich bevindt : cytoplasmatisch versus periplasmatisch versus extracellulair, en oplosbaar. of onoplosbaar. (Dit laatste aspect is niet in het voorbeeld van volgende figuur opgenomen.)")
2
IPTG-geïnduceerde productie van RP4 proteïnen : (A) DNA primase (118 en 80 kDa proteinen) ; (B) 16.5 en 8.6 kDA proteinen. Waardcelstam : E. coli HB101. Analyse door SDS-polyacrylamidegel electroforese. (A) insert in directe (c, d) of omgekeerde (e,f) orientatie ; geïnduceerd (c, e) of niet-geïnduceerd (d,f) met IPTG. (a) referentie proteïnen ; (b) gezuiverd RP4 DNA primase (118 en 80 kDa). (B) insert in directe (b, c) or of omgekeerde (d,e) orientatie ; geïnduceerd (b, d) of niet-geïnduceerd (c,e) met IPTG. (f) referentie proteïnen ; (a) gezuiverd 16.5 kDa proteïne.
 insert in directe (c, d) of omgekeerde (e,f) orientatie ; geïnduceerd (c, e) of niet-geïnduceerd (d,f) met IPTG. (a) referentie proteïnen ; (b) gezuiverd RP4 DNA primase (118 en 80 kDa). (B) insert in directe (b, c) or of omgekeerde (d,e) orientatie ; geïnduceerd (b, d) of niet-geïnduceerd (c,e) met IPTG. (f) referentie proteïnen ; (a) gezuiverd 16.5 kDa proteïne.")
3
uit : J. Sambrook & D.W. Russell : Molecular cloning, a laboratory manual. 3rd edition, 2001

4
1. Basisaspecten - regulatie - hoge expressiewaarden kunnen - toxisch zijn (door het eiwitproduct) voor de cel - "metabolic drain" veroorzaken: dus verstoring van het metabolisme => iedere verandering die het expressieniveau verlaagt (bvb. door mutaties, verlies van plasmide, ...) zal de betrokken cel een selectief groeivoordeel geven - vermijden van toxiciteit : groei de celpopulatie tot het geschikte niveau (d.w.z. aantal) vooraleer de synthese van het gekloneerde eiwit aanvangt, m.a.w. induceer de expressie (en tot dan, hou het gen onder repressie) - induceer expressie in de laat-logaritmische fase (terwijl de cellen nog voldoende metabolisch actief zijn) - regulatie is in hoofdzaak een zaak van transcriptiecontrole - andere factoren: translatie, stabiliteit (van vector, transcript en product)
 zal de betrokken. cel een selectief groeivoordeel geven. - vermijden van toxiciteit : groei de celpopulatie tot het geschikte niveau. (d.w.z. aantal) vooraleer de synthese van het gekloneerde eiwit aanvangt, m.a.w. induceer de expressie (en tot dan, hou het gen onder repressie) - induceer expressie in de laat-logaritmische fase (terwijl de cellen nog. voldoende metabolisch actief zijn) - regulatie is in hoofdzaak een zaak van transcriptiecontrole. - andere factoren: translatie, stabiliteit (van vector, transcript en product)")
5
- E.coli RNA polymerase : a2, b, b'
- transcriptie : - E.coli RNA polymerase : a2, b, b' algemene promotorstructuur (‘consensus’ sequenties) s70 promotor : TTGACA (16-19 nt) TATAAT (5-8 nt) start (+1) (zones / optimale afstanden) (17) (7) UP-elementen kunnen een extra invloed op efficientie hebben - initiatie : efficiente promotoren en hun regulatie - lacUV5 : IPTG, LacI, LacIq, titration, 'leakiness' - trp : tryptofaan + aporepressor (TrpR), IAA inductie - Ptac : upstream Plac + downstream Ptrp (cfr. PtacI, PtacII, Ptrc, en andere varianten) inductie door IPTG - PL, PR : repressor CIts857 : inductie door temperatuurshift of : CIwt + inductie by mitomycine C (nb. CIts857 is minder efficient als regulator (repressor) dan wild type CI) vervolgt
 s70 promotor : TTGACA (16-19 nt) TATAAT (5-8 nt) start (+1) (zones / optimale afstanden) -35 (17) -10 (7) UP-elementen kunnen een extra invloed op efficientie hebben. - initiatie : efficiente promotoren en hun regulatie. - lacUV5 : IPTG, LacI, LacIq, titration, leakiness - trp : tryptofaan + aporepressor (TrpR), IAA inductie. - Ptac : upstream -20 Plac + downstream -20 Ptrp. (cfr. PtacI, PtacII, Ptrc, en andere varianten) inductie door IPTG. - PL, PR : repressor CIts857 : inductie door temperatuurshift. of : CIwt + inductie by mitomycine C. (nb. CIts857 is minder efficient als regulator (repressor) dan wild type CI) vervolgt.")
6
E. coli Pribnow box (-10) T89 A99 T50 A65 A65 T100
s70 binding site (-35) T85 T83 G81 A61 C69 A52 (in feite de lacUV5 promoter ; wild-type lac promoter heeft TATGTT als -10 gebied) E. coli promoters, en hybride promoters tacI en II E. coli transcriptieterminatie (r-onafhankelijk)
 T85 T83 G81 A61 C69 A52. (in feite de lacUV5 promoter ; wild-type lac promoter heeft TATGTT als -10 gebied) E. coli promoters, en hybride promoters tacI en II. E. coli transcriptieterminatie (r-onafhankelijk)")
7
- T7 : toelevering van T7 RNA polymerase is vereist
vervolg - T7 : toelevering van T7 RNA polymerase is vereist - gekodeerd op een ander (compatibel) plasmide - gekodeerd door een l profaag (cIts) (in een E.coli lysogen) - door infectie met een M13mp-recombinant kloon van het polymerasegen (het T7 gen1) - araBAD : arabinose + AraC : positieve en negatieve regulatie => PL en PBAD worden zeer efficient geblokkeerd, Plac (en zijn derivaten) zijn ‘leaky’; met de faag T7 promotor (en andere specifieke faagpromotoren), wordt alle interferentie met het E.coli polymerase vermeden. - terminatie kan zijn : - factor-afhankelijk (Rho (r), Tau (t), NusA (van faag l)) - factor-onafhankelijk (GC-rijke stam-lusstructuur + oligoT-reeks) => meest gebruikt in expressievectoren zijn : - de fd terminator (in Ff fagen tussen gIII en gVIII, zie figuur in deel 6) - de rrnB operon terminatoren T1 en T2 - faag T7 terminator Tf
 plasmide. - gekodeerd door een l profaag (cIts) (in een E.coli lysogen) - door infectie met een M13mp-recombinant kloon van. het polymerasegen (het T7 gen1) - araBAD : arabinose + AraC : positieve en negatieve regulatie. => PL en PBAD worden zeer efficient geblokkeerd, Plac (en zijn derivaten) zijn ‘leaky’; met de faag T7 promotor (en andere specifieke faagpromotoren), wordt alle interferentie met het E.coli polymerase vermeden. - terminatie kan zijn : - factor-afhankelijk (Rho (r), Tau (t), NusA (van faag l)) - factor-onafhankelijk (GC-rijke stam-lusstructuur + oligoT-reeks) => meest gebruikt in expressievectoren zijn : - de fd terminator (in Ff fagen tussen gIII en gVIII, zie figuur in deel 6) - de rrnB operon terminatoren T1 en T2. - faag T7 terminator Tf.")
8
- sequentie motief : Shine-Dalgarno sequentie
- translatie - initiatie: RBS (ribosome binding site), AUG (of GUG, 91 vs 8% in E.coli) - sequentie motief : Shine-Dalgarno sequentie RBS > < Shine-Dalgarno (SD) motief RBS = ongeveer 55 nt tussen posities –35 en + 22 (versus AUG) SD : 16S rRNA 5'….GUACACCUCCUAOH 3' (E.coli) basispatroon : 5'….AGGA… +/- 7nt … AUG(start)… 3' efficientie is vrij onvoorspelbaar variaties : GAGG, GGAG, GGAGG - secundaire structuur - het AUG (GUG) initiatietriplet moet goed toegankelijk zijn, bvb. gelegen op de top van een stam-lusstructuur (en dus niet in baseparing betrokken) - triplets die volgen achter AUG beïnvloeden de efficientie (zoals ook de nucleotiden die het AUG voorafgaan, en uiteraard ook de secundaire structuur van dit gebied) vervolgt
, AUG (of GUG, 91 vs 8% in E.coli) - sequentie motief : Shine-Dalgarno sequentie. RBS > < Shine-Dalgarno (SD) motief. RBS = ongeveer 55 nt tussen posities –35 en (versus AUG) SD : 16S rRNA 5 ….GUACACCUCCUAOH 3 (E.coli) basispatroon : 5 ….AGGA… +/- 7nt … AUG(start)… 3 efficientie is vrij onvoorspelbaar. variaties : GAGG, GGAG, GGAGG. - secundaire structuur. - het AUG (GUG) initiatietriplet moet goed toegankelijk. zijn, bvb. gelegen op de top van een stam-lusstructuur. (en dus niet in baseparing betrokken) - triplets die volgen achter AUG beïnvloeden de efficientie. (zoals ook de nucleotiden die het AUG voorafgaan, en uiteraard ook. de secundaire structuur van dit gebied) vervolgt.")
9
- het effect van codongebruik is zeer complex
- zelfde aminozuur => multipele codons - zelfde codons => multipele tRNA's - verschillende codons => zelfde tRNA - codon-anticodon bindingssterkte - vergelijking van frequentie (in E. coli) van codongebruik tussen genen met hoog expressieniveau en deze met laag expressieniveau, toont het effect van sommige triplets (ongeacht of deze vaak gebruikt worden of niet) (zie Tabel) - met synthetische genen : kan men bewust systematisch kiezen voor telkens het 'optimale' triplet : dit garandeert een goed productierendement (hetgeen echter daarom niet het hoogst mogelijke is) - gendosiseffect - toename van aantal kopijen van de vector heeft - geen effect - een positief effect - een negatief effect (vb. expressie van het trypsinegen ) => niet voorspelbaar, geen algemene regel, kan alleen empirisch bepaald worden vervolg
 van codongebruik tussen. genen met hoog expressieniveau en deze met laag. expressieniveau, toont het effect van sommige triplets. (ongeacht of deze vaak gebruikt worden of niet) (zie Tabel) - met synthetische genen : kan men bewust systematisch kiezen. voor telkens het optimale triplet : dit garandeert een goed. productierendement (hetgeen echter daarom niet het hoogst mogelijke is) - gendosiseffect. - toename van aantal kopijen van de vector heeft. - geen effect. - een positief effect. - een negatief effect (vb. expressie van het trypsinegen ) => niet voorspelbaar, geen algemene regel, kan alleen empirisch bepaald worden. vervolg.")
10
Kodewoordgebruik in E. coli van sterk en zwak tot expressie komende genen
"sterk" : 24 genen (5253 triplets), o.a. 12 voor ribosomale proteïnen, 7 voor "outer membrane" proteïnen, en voor sommige RNA polymerase subeenheden en translatie-initiatie- en terminatiefactoren. "weak" : 18 genen (5231 triplets), o.a. voor verschillende repressorproteïnen, lac-permease, enz. kleine kadertjes : een enkel tRNA voor synonieme codons ; dus voor deze is interactiesterkte anticodon-codon bepalend
, o.a. 12 voor ribosomale proteïnen, 7 voor outer membrane proteïnen, en voor sommige RNA polymerase subeenheden en translatie-initiatie- en terminatiefactoren. weak : 18 genen (5231 triplets), o.a. voor verschillende repressorproteïnen, lac-permease, enz. kleine kadertjes : een enkel tRNA voor synonieme codons ; dus voor deze is interactiesterkte anticodon-codon bepalend.")
11
stabiliteit van het plasmide
- een transcriptieterminatiesignaal achter het doelwitgen is nodig voor stabiliteit van het plasmide - par locus (of loci) helpt voor plasmidestabiliteit (belangrijk op moment van inductie) - transcript stabiliteit : mRNA degradatie is een complex proces - zowel 5' als 3' UTR spelen een rol (bvb. een stam-lusstructuur in het 5' UTR geeft stabilisatie) - er is geen omgekeerde correlatie tussen grootte en halfwaardetijd van een mRNA bijgevolg: degradatie is niet afhankelijk van a-specifieke endonucleolytische splitsing - proteïne stabiliteit : degradatie is sterk gereguleerd in E.coli : er is een groot aantal proteasen in het cytoplasma, periplasma en aan de binnenste en buitenste membranen. vervolgt
 helpt voor plasmidestabiliteit (belangrijk op moment van inductie) - transcript stabiliteit : mRNA degradatie is een complex proces. - zowel 5 als 3 UTR spelen een rol (bvb. een stam-lusstructuur. in het 5 UTR geeft stabilisatie) - er is geen omgekeerde correlatie tussen grootte en halfwaardetijd. van een mRNA. bijgevolg: degradatie is niet afhankelijk van a-specifieke. endonucleolytische splitsing. - proteïne stabiliteit : degradatie is sterk gereguleerd in E.coli : er is een. groot aantal proteasen in het cytoplasma, periplasma. en aan de binnenste en buitenste membranen. vervolgt.")
12
N-regel van Varshavsky :
(maakte een recombinant proteïne met als N-terminaal aminozuur telkens een ander) - varianten met R, K, L, F, Y, W : halfwaardetijd (T1/2) van 2 min - varianten van de andere aminozuren (behalve P) : T1/2 = 10 uur Wat met het initiërende methionine ? : - meestal wordt het Metformyl (als eerste aminozuur van een celeigen eiwit) afgesplitst - hoe makkelijk kan dit met 'cel-vreemde' eiwitten? => niet meteen voorspelbaar - gebeurt blijkbaar makkelijkst als 2de aminozuur een korte zijketen heeft (vergemakkelijkt de splitsing door het methionine aminopeptidase) Stresscondities veroorzaken inductie van genen voor bepaalde proteasen, o.a. het protease La (lon gen) deze staan onder controle van s32 promotoren : - het gen voor deze RNA polymerase s32 subeenheid is rpoH - mutanten in rpoH kunnen een dramatische toename van expressierendement geven (en van productstabiliteit) Nog heel wat meer specifieke observaties werden gemaakt waarmee rekening kan gehouden worden. vervolg
 - varianten met R, K, L, F, Y, W : halfwaardetijd (T1/2) van 2 min. - varianten van de andere aminozuren (behalve P) : T1/2 = 10 uur. Wat met het initiërende methionine : - meestal wordt het Metformyl (als eerste aminozuur van een celeigen eiwit) afgesplitst. - hoe makkelijk kan dit met cel-vreemde eiwitten => niet meteen voorspelbaar. - gebeurt blijkbaar makkelijkst als 2de aminozuur een korte zijketen heeft. (vergemakkelijkt de splitsing door het methionine aminopeptidase) Stresscondities veroorzaken inductie van genen voor bepaalde proteasen, o.a. het protease La (lon gen) deze staan onder controle van s32 promotoren : - het gen voor deze RNA polymerase s32 subeenheid is rpoH. - mutanten in rpoH kunnen een dramatische toename van. expressierendement geven (en van productstabiliteit) Nog heel wat meer specifieke observaties werden gemaakt waarmee. rekening kan gehouden worden. vervolg.")
13
- extra regulatie (repressie) door LacI (inductie door IPTG)
2. Belangrijke expressiesystemen (inductie en regulatorische circuits) enkele voorbeelden - het pET systeem - met T7 RNA polymerase - extra regulatie (repressie) door LacI (inductie door IPTG) - toelevering van T7 RNA polymerase vanuit een aparte expressieëenheid op een l (DE3) profaag met PlacUV5 als promoter (ook onder LacI controle : lacO) - additionele controle door lysozyme van T7 LysE (or S) (gekodeerd op een (pACYC-afgeleid) compatibel plasmide) => het lysozyme bindt aan en inhibeert (residueel) T7 RNA polymerase - het binaire trp-cI systeem - dubbel systeem met compatibele plasmiden - doelwitgen stroomafwaarts van de PL promotor - synthese van CI (gekodeerd door cI repressorgen) wordt gereguleerd door Ptrp - toevoeging van tryptofaan activateert expressie van het doelwitgen door afsluiten van de CI synthese : vroeger reeds gezien : regulatie door CIts857 kan ook op basis van temperatuur, maar : die regulatie is minder strict (wt-CI bindt efficienter) én toevoeging van een inductor in grote fermentoren is eenvoudiger dan het verhogen van de temperatuur ; bovendien kan een temperatuurshift stressmechanismen (heat-shock) induceren
 enkele voorbeelden. - het pET systeem. - met T7 RNA polymerase. - extra regulatie (repressie) door LacI (inductie door IPTG) - toelevering van T7 RNA polymerase vanuit een aparte expressieëenheid op. een l (DE3) profaag met PlacUV5 als promoter (ook onder LacI controle : lacO) - additionele controle door lysozyme van T7 LysE (or S) (gekodeerd op een (pACYC-afgeleid) compatibel plasmide) => het lysozyme bindt aan en inhibeert (residueel) T7 RNA polymerase. - het binaire trp-cI systeem. - dubbel systeem met compatibele plasmiden. - doelwitgen stroomafwaarts van de PL promotor. - synthese van CI (gekodeerd door cI repressorgen) wordt gereguleerd door Ptrp. - toevoeging van tryptofaan activateert expressie van het doelwitgen door. afsluiten van de CI synthese : vroeger reeds gezien : regulatie door CIts857 kan ook. op basis van temperatuur, maar : die regulatie is minder strict (wt-CI bindt efficienter) én. toevoeging van een inductor in grote fermentoren is eenvoudiger dan het verhogen van de. temperatuur ; bovendien kan een temperatuurshift stressmechanismen (heat-shock) induceren.")
14
Het pET vectorsysteem voor proteïneproductie

15
Expressiecontrole op de Ptrp promoter door de l CI repressor

16
- dimeer AraC bindt aan I1 en O2 operatorplaatsen
- pBAD systeem - dimeer AraC bindt aan I1 en O2 operatorplaatsen (de DNA lus blokkeert transcriptie) - dimeer AraC + arabinose : bindt aan de I1 en I2 plaatsen dit is catabolietrepressie gevoelig : CAP + cAMP (cAMP = is laag als glucose concentratie hoog is) => samenspel van arabinose en glucose concentraties kan gebruikt worden als fijnregeling van het expressieniveau (nb. CAP-regulatie is ook aanwezig in het Lac operon, maar tac promotoren zijn niet CAP-afhankelijk) Operon voor synthese van AraA, AraB en AraD enzymen. Afbraak van arabinose tot D-xylulose-5-fosfaat, dat verder gemetaboliseerd wordt via de pentosefosfaatweg.
 - dimeer AraC + arabinose : bindt aan de I1 en I2 plaatsen. dit is catabolietrepressie gevoelig : CAP + cAMP. (cAMP = is laag als glucose concentratie hoog is) => samenspel van arabinose en glucose concentraties kan. gebruikt worden als fijnregeling van het expressieniveau. (nb. CAP-regulatie is ook aanwezig in het Lac operon, maar tac promotoren zijn niet CAP-afhankelijk) Operon voor synthese van AraA, AraB en AraD enzymen. Afbraak van arabinose tot D-xylulose-5-fosfaat, dat verder. gemetaboliseerd wordt via de pentosefosfaatweg.")
17
Regulatie van de PBAD promoter en expressie in pBAD vectoren

18
3. Productiestrategieën
E. coli : is het majeure "eerstelijns" organisme voor expressie => nadelen (beperkingen) - geen (of zeer weinig) post-translationele modificaties - nagenoeg geen efficiënte secretiewegen naar het medium (secretie brengt het proteïne in de periplasmatische ruimte) - extensieve S-S bruggen zijn moeilijk te vormen Waarom recombinante expressie/productie ? => opdrijven van het productierendement => zuivering vergemakkelijken => nieuwe eiwitvarianten creëren (mutaties, inserties, fusies, enz.) Het expressieproduct is : - oplosbaar of onoplosbaar - als matuur proteïne of als fusieproduct - accumuleert in cytoplasma of wordt gesecreteerd naar periplasma (type II secretie) (met E. coli is mogelijke extracellulaire secretie zeer beperkt, één voorbeeld : type I secretie van haemolysine) vervolgt
 - geen (of zeer weinig) post-translationele modificaties. - nagenoeg geen efficiënte secretiewegen naar het medium. (secretie brengt het proteïne in de periplasmatische ruimte) - extensieve S-S bruggen zijn moeilijk te vormen. Waarom recombinante expressie/productie => opdrijven van het productierendement. => zuivering vergemakkelijken. => nieuwe eiwitvarianten creëren (mutaties, inserties, fusies, enz.) Het expressieproduct is : - oplosbaar of onoplosbaar. - als matuur proteïne of als fusieproduct. - accumuleert in cytoplasma of wordt gesecreteerd naar periplasma. (type II secretie) (met E. coli is mogelijke extracellulaire secretie zeer beperkt, één voorbeeld : type I secretie van haemolysine) vervolgt.")
19
- kan aggregeren en precipiteren als 'inclusion bodies'
vervolg Expressieproduct : - als matuur proteïne - kan aggregeren en precipiteren als 'inclusion bodies' - maakt zuivering gemakkelijk - beschermt tegen afbraak - vereist achteraf solubilisatie en hervouwing => denatureer (in bvb. guanidinium hydrochloride) en (tracht te) renatureren - vertraging van synthesesnelheid kan het aggregatieproces (deels) inperken (bvb. door verlaging van de incubatietemperatuur) - het expressieconstruct vereist kritische manipulatie van het RBS gebied om efficiente (op hoog niveau) translatie-initiatie te verzekeren - het MET-probleem : wordt de initiator (formyl-)methionine verwijderd ? vervolgt
 en (tracht te) renatureren. - vertraging van synthesesnelheid kan het aggregatieproces (deels) inperken (bvb. door verlaging van de incubatietemperatuur) - het expressieconstruct vereist kritische manipulatie van het RBS gebied. om efficiente (op hoog niveau) translatie-initiatie te verzekeren. - het MET-probleem : wordt de initiator (formyl-)methionine verwijderd vervolgt.")
20
"Inclusion bodies" in E. coli
(Scanning electronmicrograph) (Thin section through the cells)
 (Thin section through the cells)")
21
Hoe de vorming van inclusion bodies vermijden (of verminderen) ?
(verbetering van de kans op eiwitvouwing) - cellen groeien bij lagere temperatuur (vertraagt het expressieproces) (of gebruik andere groeivertragende condities, bvb. samenstelling van media & pH waarden) - co-expressie van chaperonen : DnaK, GroES, GroEL - verwijdering van kritische aminozuurposities (bvb. in interferon-b) - fusie aan thioredoxine, of sommige andere eiwitten (zoals MalE)
 - cellen groeien bij lagere temperatuur (vertraagt het expressieproces) (of gebruik andere groeivertragende condities, bvb. samenstelling van media & pH waarden) - co-expressie van chaperonen : DnaK, GroES, GroEL. - verwijdering van kritische aminozuurposities (bvb. in interferon-b) - fusie aan thioredoxine, of sommige andere eiwitten (zoals MalE)")
22
- als fusieproteïne (bvb. random inserties in ScaI van cat : AGT'ACT)
vervolg - als fusieproteïne (bvb. random inserties in ScaI van cat : AGT'ACT) - N-terminale en C-terminale fusies zijn mogelijk (of multipele partners) - voordeel van C-terminale fusies : translatie-initiatieëfficiëntie wordt grotendeels behouden - fusiepartner kan doelwit zijn voor zuivering (etiket) - fusiepartner kan een activiteit hebben waarmee het productieniveau gevolgd kan worden - fusie aan een signaalpeptidesequentie kan secretie geven => secretie laat vorming van (minstens sommige) S-S bruggen toe - fusiepartner kan verankering in de membraan bewerkstellingen - een epitoop herkend door aan specifiek antilichaam kan voorzien worden (voor detectie of kwantificering of zuivering) - keuze tussen cytoplasmatische of periplasmische productie (secretie) vervolgt
 - N-terminale en C-terminale fusies zijn mogelijk (of multipele partners) - voordeel van C-terminale fusies : translatie-initiatieëfficiëntie wordt grotendeels behouden. - fusiepartner kan doelwit zijn voor zuivering (etiket) - fusiepartner kan een activiteit hebben waarmee het productieniveau. gevolgd kan worden. - fusie aan een signaalpeptidesequentie kan secretie geven. => secretie laat vorming van (minstens sommige) S-S bruggen toe. - fusiepartner kan verankering in de membraan bewerkstellingen. - een epitoop herkend door aan specifiek antilichaam kan voorzien worden. (voor detectie of kwantificering of zuivering) - keuze tussen cytoplasmatische of periplasmische productie (secretie) vervolgt.")
23
Verschillende mogelijkheden van translatiefusieconstructen
S : signaalsequentie ; I : membraanintegratiedomein ; A, B : fusiepeptiden, o.a. affiniteitsetiketten Het gebruik van membraanintegratiedomeinen laat toe het proteïne te verankeren in één van de membraansystemen of in de celwand.

24
Constitutieve expressie van het cat gen, en fusies door random klonering in de ScaI plaats
(baan 2) (banen 1, 6, 7, 9-15) Banen 4 en 5 zijn plasmide-vrije cellen ; banen 3 en 8 zijn proteïne-lengtemerkers.
 (banen 1, 6, 7, 9-15) Banen 4 en 5 zijn plasmide-vrije cellen ; banen 3 en 8 zijn proteïne-lengtemerkers.")
25
Secretie : is een bijzondere vorm van fusie
vervolg Secretie : is een bijzondere vorm van fusie - de fusiepartner (signaalpeptide) wordt verwijderd tijdens secretie - er is dus geen (extra) methionine aan de N-terminus - (de voorspelde) positie van splitsing is niet altijd gegarandeerd => meestal moet men verschillende secretiesignalen testen : van OmpA, OmpT, PelB, b-lactamase, alkalische fosfatase, enz. Sommige hiervan geven bij voldoende overexpressie ook lekkage naar het groeimedium. - !!! maar : niet alle proteinen zijn ‘secreteerbaar’ (zelfs al is er een signaalpeptide) Over het algemeen : 1) kleine peptiden : vaak stabiliteitsprobleem (afbraak van het peptide) => fusiebenadering te verkiezen : bvb. LacZ fusies 2) intermediaire grootte: weinig S-S : intracellulaire aanmaak meer S-S : periplasmische aanmaak (secretie) 3) grote proteïnen : problematischer
 wordt verwijderd tijdens secretie. - er is dus geen (extra) methionine aan de N-terminus. - (de voorspelde) positie van splitsing is niet altijd gegarandeerd. => meestal moet men verschillende secretiesignalen testen : van OmpA, OmpT, PelB, b-lactamase, alkalische fosfatase, enz. Sommige hiervan. geven bij voldoende overexpressie ook lekkage naar het groeimedium. - !!! maar : niet alle proteinen zijn ‘secreteerbaar’ (zelfs al is er een signaalpeptide) Over het algemeen : 1) kleine peptiden : vaak stabiliteitsprobleem (afbraak van het peptide) => fusiebenadering te verkiezen : bvb. LacZ fusies. 2) intermediaire grootte: weinig S-S : intracellulaire aanmaak. meer S-S : periplasmische aanmaak (secretie) 3) grote proteïnen : problematischer.")
26
4. Zuivering en verwerking
Fusies die makkelijke zuivering mogelijk maken: o.a. aan glutathion-S-transferase, MalE (MBP : maltose bindingsproteïne), oligo-His (hexa-His), enz. : partners of etiketten. Zuivering door affiniteitschromatografie of door immobilisatie gevolgd door elutie. Vrijzetting van het etiket/de partner door splitsing met specifieke proteasen zoals factor Xa, enterokinase, enz., of gebruik van ‘intein processing’. Voorbeelden van affiniteitsetiketten ("tags")
, oligo-His (hexa-His), enz. : partners of etiketten. Zuivering door. affiniteitschromatografie of door immobilisatie gevolgd door elutie. Vrijzetting van het etiket/de partner door splitsing met specifieke proteasen. zoals factor Xa, enterokinase, enz., of gebruik van ‘intein processing’. Voorbeelden van. affiniteitsetiketten ( tags )")
27
Voorbeelden van expressie-fusie systemen :
1. C-terminale fusie in pBAD/His vectoren - zes Histidine triplets - insertie in het juiste leesraam : BglII knipplaats - binding aan een kolom met geïmmobilizeerd Ni2+ - splitsingsplaats voor enterokinase (D-D-D-D-K* ) (* = splitsingsplaats) (n.b. enkele resterende aminozuren blijven aan de N-terminus na verwerking) 2. C-terminale fusie aan biotine-afhankelijke carboxylase (biotine is een essentiële cofactor voor een aantal carboxylasen) - biotine covalent aangehecht aan een lysine door biotine ligase (BirA) (E. coli heeft één gebiotinyleerd eiwit maar dit bindt in zijn natieve vorm niet aan streptavidine) het doelwit is N-terminaal als tag voorzien - immobilisatie aan streptavidine (batchgewijs of door affiniteitschromatografie) - factor Xa splitsingsplaats : I-E-G-R* of I-D-G-R* (niet gevolgd door P of R) (* = splitsingsplaats) (secundaire plaatsen meestal op G-R*)
 (* = splitsingsplaats) (n.b. enkele resterende aminozuren blijven aan de N-terminus na verwerking) 2. C-terminale fusie aan biotine-afhankelijke carboxylase. (biotine is een essentiële cofactor voor een aantal carboxylasen) - biotine covalent aangehecht aan een lysine door biotine ligase (BirA) (E. coli heeft één gebiotinyleerd eiwit maar dit bindt in zijn natieve vorm niet aan streptavidine) het doelwit is N-terminaal als tag voorzien. - immobilisatie aan streptavidine (batchgewijs of door affiniteitschromatografie) - factor Xa splitsingsplaats : I-E-G-R* of I-D-G-R* (niet gevolgd door P of R) (* = splitsingsplaats) (secundaire plaatsen meestal op G-R*)")
28
Proteïneproductie en verwerking met pBAD/His vectoren
C-terminale fusie 6 His-resten voor affiniteits- zuivering splitsing door enterokinase (D-D-D-D-K-*) na verwerking blijven nog een paar extra aminozuren aan de N-terminus
 na verwerking blijven nog. een paar extra. aminozuren aan. de N-terminus.")
29
C-terminale fusie aan biotine carboxylase
Binding op streptavidinekolommen voor zuivering. Verwerking met factor Xa protease (I-E-G-R-* of I-D-G-R-* (niet gevolgd door P of R))
)")
30
3. N-terminale fusie aan en verwerking door inteïne
(vermijdt het toevoegen van een protease bij de verwerkingsstappen) - N-terminale fusie aan inteïne + chitine bindingsdomein (CBD) => zelf-splitsing aan de N-terminus van het inteïne door een thiol - expressie vanaf de T7 promotor, in fusie met lacO => inductie door IPTG - bind het fusieproduct aan de chitinekolom - was de kolom, en equilibreer de kolom met een DTT-oplossing => in situ splitsing bij 4°C gedurende een nacht - elueer het doelwitproteïne - verwijder de fusiepartner (inteïne-CBD) van de kolom met SDS ~100 aminozuren ~50 aminozuren Splicing van een inteïne
 - N-terminale fusie aan inteïne + chitine bindingsdomein (CBD) => zelf-splitsing aan de N-terminus van het inteïne door een thiol. - expressie vanaf de T7 promotor, in fusie met lacO. => inductie door IPTG. - bind het fusieproduct aan de chitinekolom. - was de kolom, en equilibreer de kolom met een DTT-oplossing. => in situ splitsing bij 4°C gedurende een nacht. - elueer het doelwitproteïne. - verwijder de fusiepartner (inteïne-CBD) van de kolom met SDS. ~100 aminozuren. ~50 aminozuren. Splicing van een inteïne.")
31
Verwerking van een N-terminale fusies aan inteïne, met immobilisatie op een chitinekolom

32
4. MalE-fusies : vectoren pMAL-c2 en pMAL-p2
(met (c) of zonder (p) de signaalpeptidesequentie van het malE gen : cytoplasmische of periplasmatische expressie) - expressie-eenheid tussen de Ptac promotor en rrnB terminatoren, met lacIq op de vector om voldoende Lac-repressor te voorzien. - fusie tussen de malE partner en lacZa, gescheiden door 10xD en een MCS voor insertie van het doelwitgen. De in-frame constructie produceert blauwe kolonies op BCIG-media. Het polaire 10xD peptide scheidt de twee proteïnedelen (eigen vouwing). Klonering in de MCS geeft witte (kleurloze) kolonies. - affiniteitszuivering op een amylosekolom : na elutie met maltose en splitsing door factor Xa, wordt fusiepartner MalE verwijderd door een 2e chromatografie op amylose. - MCS bevat (aan de linkerzijde) een XmnI, die overlapt met de factor Xa knipplaats en een exacte fusie aan het doelwitgen mogelijk maakt : andere insertieplaatsen laten enkele extra aminozuren aan het N-terminale uiteinde na factor Xa splitsing. - de EcoRI kloneerplaats ligt in hetzelfde leesraam als de EcoRI plaats in lacZ : dit vergemakkelijkt de uitwisseling van een gen dat als een expressiefusie gekloneerd is in (ondermeer) de vektor lgt11.
 of zonder (p) de signaalpeptidesequentie van het malE gen : cytoplasmische of periplasmatische expressie) - expressie-eenheid tussen de Ptac promotor en rrnB terminatoren, met lacIq. op de vector om voldoende Lac-repressor te voorzien. - fusie tussen de malE partner en lacZa, gescheiden door 10xD en een MCS. voor insertie van het doelwitgen. De in-frame constructie produceert blauwe kolonies op BCIG-media. Het polaire 10xD peptide scheidt de twee proteïnedelen (eigen vouwing). Klonering in de MCS geeft witte (kleurloze) kolonies. - affiniteitszuivering op een amylosekolom : na elutie met maltose en splitsing door. factor Xa, wordt fusiepartner MalE verwijderd door een 2e chromatografie op amylose. - MCS bevat (aan de linkerzijde) een XmnI, die overlapt met de factor Xa knipplaats. en een exacte fusie aan het doelwitgen mogelijk maakt : andere insertieplaatsen. laten enkele extra aminozuren aan het N-terminale uiteinde na factor Xa splitsing. - de EcoRI kloneerplaats ligt in hetzelfde leesraam als de EcoRI plaats in. lacZ : dit vergemakkelijkt de uitwisseling van een gen dat als een. expressiefusie gekloneerd is in (ondermeer) de vektor lgt11.")
33
pMAL expressievectoren
malE : kodeert voor maltosebindend proteïne (MBP)
")
34
Karakteristieken van pMAL vectoren :
ColE1 ori ; M13 ori ; Ptac ; rrnB terminators ; lacIq ; bla Fusieconstruct : malE - DDDDDDDDDD - IEGR - MCS(polylinker) - lacZa (bij doorlezing in het leesraam : blauwkleuring met IPTG + BCIG) pMAL-c2 : malE signaalpeptidesequentie is gedeleteerd pMAL-p2 : met malE signaalpeptidesequentie : secretie naar het periplasma fusie in XmnI : exacte koppeling van het doelwitproteïne aan de factor Xa sequentie 10 Asp eenheden (D) scheiden de twee fusiegedeelten insertie in EcoRI is identisch (qua leesraam) als in lgt11 (lacZ) Splitsingsplaats van factor Xa I - E - G - R * XmnI nnn nnn nGA Ann nnT TCn nnn ATC-GAG-GGA-AGG-ATT-TCA-GAA-TTC- EcoRI
 - lacZa. (bij doorlezing in het leesraam : blauwkleuring met IPTG + BCIG) pMAL-c2 : malE signaalpeptidesequentie is gedeleteerd. pMAL-p2 : met malE signaalpeptidesequentie : secretie naar het periplasma. fusie in XmnI : exacte koppeling van het doelwitproteïne aan de factor Xa sequentie. 10 Asp eenheden (D) scheiden de twee fusiegedeelten. insertie in EcoRI is identisch (qua leesraam) als in lgt11 (lacZ) Splitsingsplaats van factor Xa I - E - G - R * XmnI nnn nnn nGA Ann nnT TCn nnn. ATC-GAG-GGA-AGG-ATT-TCA-GAA-TTC- EcoRI.")
35
"Multiple cloning site" in (één van) de pMAL vectoren
Andere mogelijkheid (in sommige vectoren) Pro-Gly-Ala-Ala-His-Tyr : splitsing door Genenase I (een geëngineerde vorm van subtilisine) splitst achter His-Tyr (afhankelijke van het volgende aminozuur) (splitst His-Tyr-Glu en His-Tyr-Asp traag, en His-Tyr-Pro en His-Tyr-Ile niet)
 Pro-Gly-Ala-Ala-His-Tyr : splitsing door Genenase I (een geëngineerde vorm van subtilisine) splitst achter His-Tyr (afhankelijke van het volgende aminozuur) (splitst His-Tyr-Glu en His-Tyr-Asp traag, en His-Tyr-Pro en His-Tyr-Ile niet)")
36
Verwerking van expressieproducten uit fusieconstructen in pMAL vectoren

37
- gelijkaardige elementen als in vorige voorbeelden :
5. Pinpoint vectoren - gelijkaardige elementen als in vorige voorbeelden : tac (en T7) promoter, insertieplaatsen, etiket, factor Xa doelwit, enz. hier tevens SP6 (naast T7 vanop andere streng) promoter voor synthese van RNA - N-teminaal is een etiket waarin een lysine gebiotinyleerd wordt door het E. coli biotin ligase holoenzyme Het lysine is toegankelijk voor (strept)avidine : => etiket voor detectie en zuivering E. coli produceert slechts één enkel endogeen gebiotinyleerd proteïne dat, in zijn natieve conformatie, niet bindt aan (strept)avidine : => zeer specifiek zuiveringssysteem voor het recombinante fusieproteïne. Ook, detectie van het fusieproteïne mogelijk met streptavidine-alkalische fosfatase. - het fusieproteïne bindt aan (strept)avidine, en kan geëlueerd worden door toevoeging van 5mM biotin : => geen denaturaterende condities vereist.
 promoter, insertieplaatsen, etiket, factor Xa doelwit, enz. hier tevens SP6 (naast T7 vanop andere streng) promoter voor synthese van RNA. - N-teminaal is een etiket waarin een lysine gebiotinyleerd wordt door het. E. coli biotin ligase holoenzyme. Het lysine is toegankelijk voor (strept)avidine : => etiket voor detectie en zuivering. E. coli produceert slechts één enkel endogeen gebiotinyleerd proteïne dat, in zijn natieve. conformatie, niet bindt aan (strept)avidine : => zeer specifiek zuiveringssysteem voor het. recombinante fusieproteïne. Ook, detectie van het fusieproteïne mogelijk met streptavidine-alkalische fosfatase. - het fusieproteïne bindt aan (strept)avidine, en kan geëlueerd worden door. toevoeging van 5mM biotin : => geen denaturaterende condities vereist.")
38
twee promoters : T7 en tac factor Xa knipplaats
(Commerciële) Pinpoint vectoren twee promoters : T7 en tac factor Xa knipplaats N-terminale biotinylering (tag) polylinker (MCS) in vitro RNA sondes met de SP6 (of T7) promoter secretiesignaal (sommige vectoren) Detectie door streptavidine-alkalische fosfatase Elutie van fusieproteïne met 5 mM biotine (dus : geen denaturatie vereist)
 Pinpoint vectoren. twee promoters : T7 en tac. factor Xa knipplaats. N-terminale biotinylering (tag) polylinker (MCS) in vitro RNA sondes met de. SP6 (of T7) promoter. secretiesignaal (sommige vectoren) Detectie door. streptavidine-alkalische. fosfatase. Elutie van fusieproteïne met. 5 mM biotine. (dus : geen denaturatie vereist)")
39
Recenter voorbeeld met TOPO-klonering en gebruik van TEV protease bij
verwerking TEV (van Tobacco etch virus) protease : herkent E-X-X-Y-X-Q-G/S splitst tussen Q en G of S Tolereert heel wat aminozuursequentie maar de splitsingsefficientie varieert sterk. Meestal wordt E-N-L-Y-F-Q-S gebruikt dat het meest efficientie substraat lijkt. Structuur similair aan serineproteasen maar met C i.p.v. S in de katalytische triade Ser-Asp-His, hetgeen allicht zijn resistentie tegen veel protease-inhibitoren verklaart.
 protease : herkent E-X-X-Y-X-Q-G/S. splitst tussen Q en G of S. Tolereert heel wat aminozuursequentie. maar de splitsingsefficientie varieert sterk. Meestal wordt E-N-L-Y-F-Q-S gebruikt. dat het meest efficientie substraat lijkt. Structuur similair aan serineproteasen maar. met C i.p.v. S in de katalytische triade. Ser-Asp-His, hetgeen allicht zijn resistentie. tegen veel protease-inhibitoren verklaart.")
40
Recenter voorbeeld met
sacB als selectiemerker en LIC klonering. TEV protease voor verwerking. Fasmide vector.

41
5. Optimalisatiestrategieën
bvb. C-terminale fusie aan een reportergen om expressierendement te testen in functie van (kleine) wijzigingen. Ondermeer door regio-specifieke willekeurige mutaties (o.a. in het gebied van ribosoombinding en initiatie) kan de beste producer geïdentificeerd worden. Op analoge wijze kan een N-terminale fusie aan een reportergen voorzien worden, en dusdanig dat na optimalisatie (regio-mutagenese) kan het reportergen geëxciseerd kan worden. Er wordt dan op gespeculeerd dat het C-terminale deel van de fusie geen invloed heeft op de expressieëfficientie.
 wijzigingen. Ondermeer door regio-specifieke. willekeurige mutaties (o.a. in het gebied van ribosoombinding en initiatie) kan de. beste producer geïdentificeerd worden. Op analoge wijze kan een N-terminale fusie aan een reportergen voorzien. worden, en dusdanig dat na optimalisatie (regio-mutagenese) kan het. reportergen geëxciseerd kan worden. Er wordt dan op gespeculeerd. dat het C-terminale deel van de fusie geen invloed heeft op de. expressieëfficientie.")
42
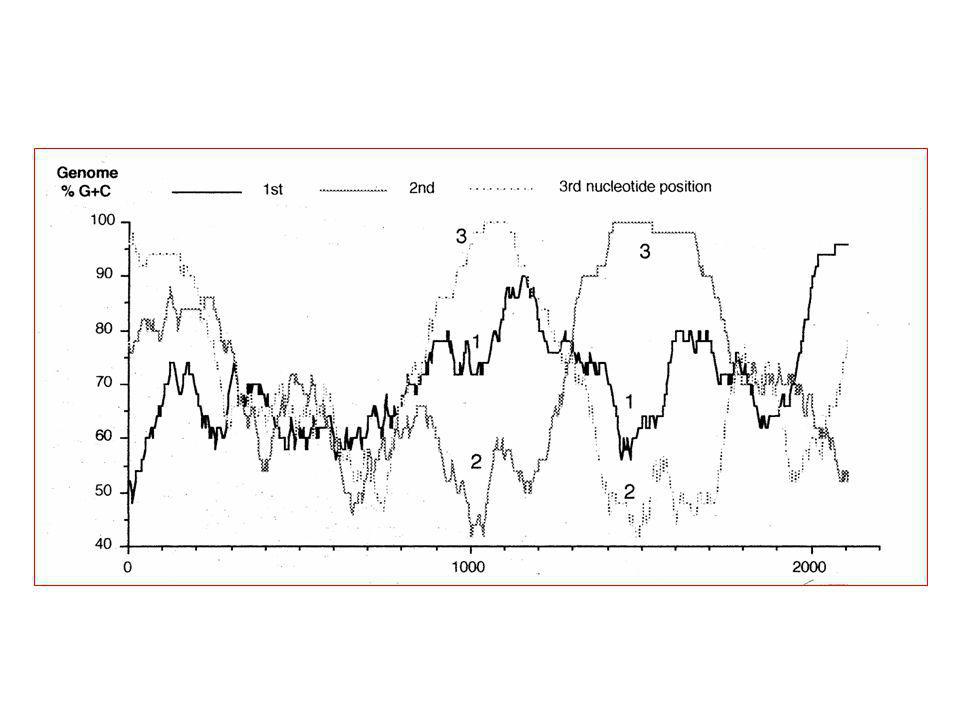
Addendum : Genoomstrategie : lokaliseren van coderende gebieden (ORFs) in een genoomsequentie
 in een genoomsequentie")
Verwante presentaties