Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Populaties

2
Populatie-ecologie Populatie. Groep van individuen van eenzelfde soort die op eenzelfde tijdstip op een welbepaalde plaats leven en met elkaar interageren.

3
Populatie-ecologie Populatie. Groep van individuen van eenzelfde soort die op eenzelfde tijdstip op een welbepaalde plaats leven en met elkaar interageren. Vertoont eigenschappen die we niet aantreffen bij de individuele organismen waaruit ze is samengesteld.

4
Populatie-ecologie Populatie. Groep van individuen van eenzelfde soort die op eenzelfde tijdstip op een welbepaalde plaats leven en met elkaar interageren. Vertoont eigenschappen die we niet aantreffen bij de individuele organismen waaruit ze is samengesteld. Wordt gekenmerkt door gemeenschappelijk genetisch reservoir en vormt eenheid waarop natuurlijke selectie via veranderingen in allelfrekwenties inwerkt.

5
Populatie-ecologie Populatie. Groep van individuen van eenzelfde soort die op eenzelfde tijdstip op een welbepaalde plaats leven en met elkaar interageren. Vertoont eigenschappen die we niet aantreffen bij de individuele organismen waaruit ze is samengesteld. Wordt gekenmerkt door gemeenschappelijk genetisch reservoir en vormt eenheid waarop natuurlijke selectie via veranderingen in allelfrekwenties inwerkt. Grote belangstelling vanuit toegepaste invalshoeken zoals bosbouw, agronomie, pestcontrole, wildbeheer en natuurbeheer.

6
Densiteit

7
Densiteit Begrip ‘populatiegrootte’ heeft enkel betekenis indien grenzen van populatie (of steekproef ervan) bekend zijn. Uitgedrukt als densiteit (= aantal individuen per oppervlakte/volume eenheid).
 bekend zijn. Uitgedrukt als densiteit (= aantal individuen per oppervlakte/volume eenheid).")
8
Densiteit Begrip ‘populatiegrootte’ heeft enkel betekenis indien grenzen van populatie (of steekproef ervan) bekend zijn. Uitgedrukt als densiteit (= aantal individuen per oppervlakte/volume eenheid).
 bekend zijn. Uitgedrukt als densiteit (= aantal individuen per oppervlakte/volume eenheid).")
9
Densiteit Densiteitsbepalingen gebeuren meestal aan de hand van random steekproeven (sampling).
.")
10
Densiteit Densiteitsbepalingen gebeuren meestal aan de hand van random steekproeven (sampling). Nauwkeurigheid neemt toe met aantal steekproeven (studieplots) en homogeniteit van bemonsterde habi-tat.
 en homogeniteit van bemonsterde habi-tat.")
11
Densiteit Densiteitsbepalingen gebeuren meestal aan de hand van random steekproeven (sampling). Nauwkeurigheid neemt toe met aantal steekproeven (studieplots) en homogeniteit van bemonsterde habi-tat. Merk-hervangst (mark-recapture) binnen afgebakende grenzen van een studiegebied.
 en homogeniteit van bemonsterde habi-tat. Merk-hervangst (mark-recapture) binnen afgebakende grenzen van een studiegebied.")
12
Densiteit

13
Densiteit Proportie gemerkte (en gevangen) individuen in tweede vangstsessie gelijk aan proportie gemerkte individuen in volledige populatie.

14
Densiteit Ngemerkte individuen in vangst 1 x Nindividuen in vangst 2
Proportie gemerkte (en gevangen) individuen in tweede vangstsessie gelijk aan proportie gemerkte individuen in volledige populatie. Ngemerkte individuen in vangst 1 x Nindividuen in vangst 2 Nhervangsten in vangst 2 Ntot =
 individuen in tweede vangstsessie gelijk aan proportie gemerkte individuen in volledige populatie. Ngemerkte individuen in vangst 1 x Nindividuen in vangst 2 Nhervangsten in vangst 2. Ntot =")
15
Ruimtelijke distributie

16
Ruimtelijke distributie
Geaggregeerde distributie (clumped, patchy). Indivi-duen zijn geconcentreerd in welbepaalde delen van het habitat.
. Indivi-duen zijn geconcentreerd in welbepaalde delen van het habitat.")
17
Ruimtelijke distributie
Geaggregeerde distributie (clumped, patchy). Indivi-duen zijn geconcentreerd in welbepaalde delen van het habitat.
. Indivi-duen zijn geconcentreerd in welbepaalde delen van het habitat.")
18
Ruimtelijke distributie
Uniforme distributie. Meer gelijkmatige spreiding dan verwacht op basis van willekeurig (random) gebruik.
 gebruik.")
19
Ruimtelijke distributie
Uniforme distributie. Meer gelijkmatige spreiding dan verwacht op basis van willekeurig (random) gebruik.
 gebruik.")
20
Ruimtelijke distributie
Willekeurige distributie. Distributie van individuen is niet gerelateerd aan die van andere individuen.

21
Ruimtelijke distributie
Willekeurige distributie. Distributie van individuen is niet gerelateerd aan die van andere individuen.

22
Abundantie versus distributie

23
Abundantie versus distributie
Kernvraag. Zijn wijdverbreide soorten abundant en hebben zeldzame soorten een kleine verspreiding ? (Herbert Andrewartha en Charles Birch)
")
24
Abundantie versus distributie
Kernvraag. Zijn wijdverbreide soorten abundant en hebben zeldzame soorten een kleine verspreiding ?

25
Abundantie versus distributie
Hypothesen Zeldzame soorten zijn moeilijker detecteerbaar.

26
Abundantie versus distributie
Hypothesen Zeldzame soorten zijn moeilijker detecteerbaar. Generalisten zijn meer algemeen en wijdverbreid (ecologische populatiemodel).
.")
27
Abundantie versus distributie
Hypothesen Zeldzame soorten zijn moeilijker detecteerbaar. Generalisten zijn meer algemeen en wijdverbreid (ecologische populatiemodel). Minder mobiele soorten bezetten minder habitat- vlekken (lokaal populatiemodel).
. Minder mobiele soorten bezetten minder habitat- vlekken (lokaal populatiemodel).")
28
parameter schatter 838 ± 341 95% CI 240 – 1591
Abundantie versus distributie mobiliteit parameter schatter 838 ± % CI 240 – 1591 bezettingsgraad

29
Habitat selectie

30
Habitat selectie Habitat selectie. Organismen verkiezen bepaalde compo-nenten van het habitat boven andere.

31
Habitat selectie Habitat selectie. Organismen verkiezen bepaalde compo-nenten van het habitat boven andere. Kan zowel proximaat als ultimaat (evolutionair) bena-derd worden.
 bena-derd worden.")
32
Habitat selectie Habitat selectie. Organismen verkiezen bepaalde compo-nenten van het habitat boven andere. Kan zowel proximaat als ultimaat (evolutionair) bena-derd worden. Ideal free distribution model van S.D. Fretwell (1972).
 bena-derd worden. Ideal free distribution model van S.D. Fretwell (1972).")
33
Habitat selectie

34
Habitat selectie

35
Habitat selectie Habitat selectie. Organismen verkiezen bepaalde compo-nenten van het habitat boven andere. Kan zowel proximaat als ultimaat (evolutionair) bena-derd worden. Ideal free distribution model van Steve Fretwell (1972). Ideal despotic distribution model (Fretwell)
 bena-derd worden. Ideal free distribution model van Steve Fretwell (1972). Ideal despotic distribution model (Fretwell)")
36
Demografie

37
Demografie Soorten verschillen in de manier waarop energie geïnvesteerd wordt in voortplanting.

38
Demografie Soorten verschillen in de manier waarop energie geïnvesteerd wordt in voortplanting. Semelpaar. Eénmalige en zeer grote investering.

39
Demografie Soorten verschillen in de manier waarop energie geïnvesteerd wordt in voortplanting. Semelpaar. Eénmalige en zeer grote investering. Iteropaar. Herhaalde en kleinere investeringen.

40
Demografie Soorten verschillen in de manier waarop energie geïnvesteerd wordt in voortplanting. Semelpaar. Eénmalige en zeer grote investering. Iteropaar. Herhaalde en kleinere investeringen. Vormt een belangrijk kenmerk van de levensgeschiedenis (life history trait) van organismen.
 van organismen.")
41
Demografie r-selectie. Verwijst naar aanwezigheid van levens-kenmerken die bijdragen tot hoge populatiegroei.

42
Demografie K-selectie. Verwijst naar aanwezigheid van levens-kenmerken die bijdragen tot hoge overleving.

43
Overleving Mortaliteits- en overlevingsparameters worden weerge-geven in levens- of sterftetabellen (life tables).
.")
44
Overleving Mortaliteits- en overlevingsparameters worden weerge-geven in levens- of sterftetabellen (life tables). Laat toe de kans te berekenen dat een individu van een welbepaalde cohorte overleeft tot een welbepaalde leeftijd.

45
Overleving

46
Overleving

47
Demografie Mortaliteits- en overlevingsparameters worden weerge-geven in zogenaamde levens- of sterftetabellen (life tables). Laat toe de kans te berekenen dat een individu van een welbepaalde cohorte overleeft tot een welbepaalde leeftijd. Leeftijdsstructuur verwijst naar het aantal en de proportie individuen van elk geslacht in elke leeftijds-categorie.

48
Demografie

49
Populatiegroei

50
Populatiegroei Aangezien geen enkele populatie oneindig in aantal toe-neemt, wordt de groei van populaties blijkbaar gereguleerd.

51
Populatiegroei Aangezien geen enkele populatie oneindig in aantal toe-neemt, wordt de groei van populaties blijkbaar gereguleerd. Groei of afname in populatiegrootte (per eenheid oppervlakte) is voornamelijk het resultaat van nataliteit (gemiddelde per capita geboorte-ratio) en mortaliteit (gemiddelde per capita sterfte-ratio).
 is voornamelijk het resultaat van nataliteit (gemiddelde per capita geboorte-ratio) en mortaliteit (gemiddelde per capita sterfte-ratio).")
52
Populatiegroei Aangezien geen enkele populatie oneindig in aantal toe-neemt, wordt de groei van populaties blijkbaar gereguleerd. Groei of afname in populatiegrootte (per eenheid oppervlakte) is voornamelijk het resultaat van nataliteit (gemiddelde per capita geboorte-ratio) en mortaliteit (gemiddelde per capita sterfte-ratio). Om de ratio van ‘verandering’ in populatie-grootte te kennen, is tevens kennis van een tijdsinterval vereist.
 is voornamelijk het resultaat van nataliteit (gemiddelde per capita geboorte-ratio) en mortaliteit (gemiddelde per capita sterfte-ratio). Om de ratio van ‘verandering’ in populatie-grootte te kennen, is tevens kennis van een tijdsinterval vereist.")
53
Populatiegroei ∆N/∆t = N (b - d)
")
54
Populatiegroei ∆N/∆t = N (b - d)
De groeiratio op een willekeurig moment in de tijd bedraagt dan: dN/dt = rN

55
Populatiegroei

56
Populatiegroei ∆N/∆t = N (b - d)
De groeiratio op een willekeurig moment in de tijd bedraagt dan: dN/dt = rN Intrinsieke groeiratio (rmax): maximum ratio waarmee een populatie in aantal kan toenemen onder ideale om-standigheden.
: maximum ratio waarmee een populatie in aantal kan toenemen onder ideale om-standigheden.")
57
Populatiegroei Factoren zoals predatie, competitie, schaarste aan kritische hulpbronnen, etc. verhinderen een ongelimi-teerde exponentiële toename in populatiegrootte. Draagkracht (K, carrying capacity): maximale populatie-grootte onder gegeven milieuomstandigheden (r = 0).
: maximale populatie-grootte onder gegeven milieuomstandigheden (r = 0).")
58
Populatiegroei Factoren zoals predatie, competitie, schaarste aan kritische hulpbronnen, etc. verhinderen een ongelimi-teerde exponentiële toename in populatiegrootte. Draagkracht (K, carrying capacity): maximale populatie-grootte onder gegeven milieuomstandigheden (r = 0). Logistische vergelijking: dN/dt = rN(K –N)/K
: maximale populatie-grootte onder gegeven milieuomstandigheden (r = 0). Logistische vergelijking: dN/dt = rN(K –N)/K")
59
Populatiegroei

60
Populatiegroei Zowel de exponentiële als logistische groeicurve vormen een sterke oversimplificatie van groeipatronen in natuur-lijke populaties.

61
Populatiegroei Zowel de exponentiële als logistische groeicurve vormen een sterke oversimplificatie van groeipatronen in natuur-lijke populaties. oscillaties rond K

62
Populatiegroei Zowel de exponentiële als logistische groeicurve vormen een sterke oversimplificatie van groeipatronen in natuur-lijke populaties. oscillaties rond K time lag modellen

63
Populatiegroei

64
Populatiegroei Zowel de exponentiële als logistische groeicurve vormen een sterke oversimplificatie van groeipatronen in natuur-lijke populaties. oscillaties rond K time lag modellen stochastische modellen

65
Populatiegroei

66
Populatieregulatie

67
Populatieregulatie Historische discussies omwille van onduidelijke definiëring van de termen limiterende en regulerende factoren.

68
Populatieregulatie Historische discussies omwille van onduidelijke definiëring van de termen limiterende en regulerende factoren. Limiterende factor: verandering in gemiddelde of evenwichtsdensiteit.

69
Populatieregulatie Historische discussies omwille van onduidelijke definiëring van de termen limiterende en regulerende factoren. Limiterende factor: verandering in gemiddelde of evenwichtsdensiteit. Regulerende factor: verhoogde mortaliteit (of verlaagde reproductie) bij toenemende densiteit (proportioneel effect)
 bij toenemende densiteit (proportioneel effect)")
70
Populatieregulatie Historische discussies omwille van onduidelijke definiëring van de termen limiterende en regulerende factoren. Limiterende factor: verandering in gemiddelde of evenwichtsdensiteit. Regulerende factor: verhoogde mortaliteit (of verlaagde reproductie) bij toenemende densiteit (proportioneel effect) Onenigheid omtrent relatieve belang van densiteits-afhankelijke versus -onafhankelijke factoren.
 bij toenemende densiteit (proportioneel effect) Onenigheid omtrent relatieve belang van densiteits-afhankelijke versus -onafhankelijke factoren.")
71
Populatieregulatie predatie, ziekte, competitie, …

72
Stephens P.A. et al. 1999. Oikos 87: 185-190

73
Populatiedynamiek

74
Populatiedynamiek Populatie-ecologen bestuderen de dynamiek van natuur-lijke populaties via key-factor analyse en experimenten.

75
Populatiedynamiek Populatie-ecologen bestuderen de dynamiek van natuur-lijke populaties via key-factor analyse en experimenten. Key-factor analyse Restrospectieve analyse van jaarlijkse veranderingen in mortaliteit en reproductie via opstellen van levenstabel-len met discrete levensstadia.

76
Populatiedynamiek

77
Populatiedynamiek ∑ k = loge(Ns) – loge(Ne)
 – loge(Ne)")
78
Populatiedynamiek

79
Populatiedynamiek

80
Populatiedynamiek Populatie-ecologen bestuderen de dynamiek van natuur-lijke populaties via key-factor analyse en experimenten. Experimentele studie Identificatie van limiterende factoren en experimentele studie van hun impact op populatie fluctuaties.

81
Populatiedynamiek D. Spiller & T. Schoener, University of California

82
Populatiedynamiek

83
Toegepaste populatie-ecologie

84
Toegepaste populatie-ecologie
Populatie-ecologie levert belangrijke inzichten voor het duurzaam (commercieel) beheer van natuurlijke popula-ties.
 beheer van natuurlijke popula-ties.")
85
Toegepaste populatie-ecologie
Populatie-ecologie levert belangrijke inzichten voor het duurzaam (commercieel) beheer van natuurlijke popula-ties. Hoe groot kan yield zijn zonder gevaar voor voortbestaan op lange termijn ?
 beheer van natuurlijke popula-ties. Hoe groot kan yield zijn zonder gevaar voor voortbestaan op lange termijn")
86
Toegepaste populatie-ecologie
E.S. Russell (1931). Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst.
. Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst.")
87
Toegepaste populatie-ecologie
E.S. Russell (1931). Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst. S2 = S1 + R + G - M - F
. Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst. S2 = S1 + R + G - M - F.")
88
Toegepaste populatie-ecologie
E.S. Russell (1931). Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst. S2 = S1 + R + G - M - F Duurzame bevissing R + G = M + F
. Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst. S2 = S1 + R + G - M - F. Duurzame bevissing R + G = M + F.")
89
Toegepaste populatie-ecologie
E.S. Russell (1931). Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst. S2 = S1 + R + G - M - F Duurzame bevissing R + G = M + F Na aanvang van bevissing wordt populatiegrootte gere-duceerd waarbij F densiteitsafhankelijk kan gecompen-seerd worden door een hogere R of snellere G of lagere M.
. Oogstbare gedeelte van visbestand (stock uitgedrukt in gewicht) verhoogt t.g.v. reproductie, immigratie en groei en verlaagt t.g.v. natuurlijke morta-liteit en vangst. S2 = S1 + R + G - M - F. Duurzame bevissing R + G = M + F. Na aanvang van bevissing wordt populatiegrootte gere-duceerd waarbij F densiteitsafhankelijk kan gecompen-seerd worden door een hogere R of snellere G of lagere M.")
90
Toegepaste populatie-ecologie
Op welk niveau van stabilisatie (R+G=M+F) is F maximaal ?
 is F maximaal")
91
Toegepaste populatie-ecologie
Op welk niveau van stabilisatie (R+G=M+F) is F maximaal ? Sigmoïd-curve theorie: een maximale yield wordt beko-men door een populatie rond de helft van haar draagkracht (K) te houden.
 is F maximaal Sigmoïd-curve theorie: een maximale yield wordt beko-men door een populatie rond de helft van haar draagkracht (K) te houden.")
92
Toegepaste populatie-ecologie
dN/dt = rN(K –N)/K
/K")
93
Ruimtelijk gestructureerde populaties

94
Ruimtelijk gestructureerde populaties
Onder natuurlijke omstandigheden komen soorten voor als sets van afzonderlijke, ruimtelijk gescheiden deelpopula-ties.

95
Ruimtelijk gestructureerde populaties
Onder natuurlijke omstandigheden komen soorten voor als sets van afzonderlijke, ruimtelijk gescheiden deelpopula-ties.

96
Ruimtelijk gestructureerde populaties
Onder natuurlijke omstandigheden komen soorten voor als sets van afzonderlijke, ruimtelijk gescheiden deelpopula-ties.

97
Ruimtelijk gestructureerde populaties
Onder natuurlijke omstandigheden komen soorten voor als sets van afzonderlijke, ruimtelijk gescheiden deelpopula-ties. Elke deelpopulatie wordt gekenmerkt door specifieke demografische parameters.

98
Ruimtelijk gestructureerde populaties
Onder natuurlijke omstandigheden komen soorten voor als sets van afzonderlijke, ruimtelijk gescheiden deelpopula-ties. Elke deelpopulatie wordt gekenmerkt door specifieke demografische parameters. Bij metapopulaties heeft ook de uitwisseling van indivi-duen tussen deelpopulaties een belangrijke demografische impact.

99
Metapopulatie theorie
Metapopulaties verwijzen naar ruimtelijk opgedeelde po-pulaties waarvan de dynamiek op twee verschillende niveaus werkt:

100
Metapopulatie theorie
Metapopulaties verwijzen naar ruimtelijk opgedeelde po-pulaties waarvan de dynamiek op twee verschillende niveaus werkt: binnen deelpopulaties: geboorte, sterfte, immigratie en emigratie.

101
Metapopulatie theorie
Metapopulaties verwijzen naar ruimtelijk opgedeelde po-pulaties waarvan de dynamiek op twee verschillende niveaus werkt: binnen deelpopulaties: geboorte, sterfte, immigratie en emigratie. tussen deelpopulaties: kolonisatie en extinctie.

102
Metapopulatie theorie

103
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson).
.")
104
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson).
.")
105
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson). Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten.
. Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten.")
106
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson). Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten.
. Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten.")
107
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson). Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten. N=0
. Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten. N=0.")
108
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson). Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten. N=K
. Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten. N=K.")
109
Metapopulatie theorie
Metapopulatie theorie werd ontwikkeld op het einde van jaren ’60 ongeveer gelijktijdig met theorie van eiland-biogeografie (MacArthur & Wilson). Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten. Laat voorspellingen toe omtrent de proportie bezette habitatvlekken als functie van extinctie en kolonisatie.
. Levins’ metapopulatie model beschrijft een oneindig aantal habitatplekken die elk al dan niet een lokale populatie van de beschouwde soort bevatten. Laat voorspellingen toe omtrent de proportie bezette habitatvlekken als functie van extinctie en kolonisatie.")
110
Metapopulatie theorie
dp/dt = k.p(1 - p) - e.p p = proportie habitatvlekken bezet op tijdstip t e = extinctiekans k = kolonisatiekans
 - e.p. p = proportie habitatvlekken bezet op tijdstip t. e = extinctiekans. k = kolonisatiekans.")
111
Metapopulatie theorie
dp/dt = k.p(1 - p) - e.p dp/dt = (k - e).p.[1 - p/(1 - e/k)]
 - e.p. dp/dt = (k - e).p.[1 - p/(1 - e/k)]")
112
Metapopulatie theorie
dp/dt = k.p(1 - p) - e.p = k.p - k.p² - e.p = k.p - p(k.p) - e.p = (k-e)p - p(k.p) = (k-e)p.1 -((p.k)/k-e) = (k-e)p.1-(p/((k-e)/k)) dp/dt = (k - e).p.[1 - p/(1 - e/k)]
 - e.p. = k.p - k.p² - e.p. = k.p - p(k.p) - e.p. = (k-e)p - p(k.p) = (k-e)p.1 -((p.k)/k-e) = (k-e)p.1-(p/((k-e)/k)) dp/dt = (k - e).p.[1 - p/(1 - e/k)]")
113
Metapopulatie theorie
dp/dt = k.p(1 - p) - e.p = k.p - k.p² - e.p = k.p - p(k.p) - e.p = (k-e)p - p(k.p) = (k-e)p.1 -((p.k)/k-e) = (k-e)p.1-(p/((k-e)/k)) dp/dt = (k - e).p.[1 - p/(1 - e/k)] dN/dt = r.N.[(K- N)/K]
 - e.p. = k.p - k.p² - e.p. = k.p - p(k.p) - e.p. = (k-e)p - p(k.p) = (k-e)p.1 -((p.k)/k-e) = (k-e)p.1-(p/((k-e)/k)) dp/dt = (k - e).p.[1 - p/(1 - e/k)] dN/dt = r.N.[(K- N)/K]")
114
Metapopulatie theorie
K = 1 - e/k Tijd Proportie bezette vlekken dp/dt = k.p(1 - p) - e.p = k.p - k.p² - e.p = k.p - p(k.p) - e.p = (k-e)p - p(k.p) = (k-e)p.1 -((p.k)/k-e) = (k-e)p.1-(p/((k-e)/k)) dN/dt = r.N.[(K- N)/K]
 - e.p. = k.p - k.p² - e.p. = k.p - p(k.p) - e.p. = (k-e)p - p(k.p) = (k-e)p.1 -((p.k)/k-e) = (k-e)p.1-(p/((k-e)/k)) dN/dt = r.N.[(K- N)/K]")
115
Metapopulatie theorie
Complexere modellen houden rekening met variatie in vlekgrootte

116
Metapopulatie theorie
Complexere modellen houden rekening met variatie in vlekgrootte vlekkwaliteit

117
Metapopulatie theorie
Complexere modellen houden rekening met variatie in vlekgrootte vlekkwaliteit ruimtelijke configuratie

118
Metapopulatie theorie
Complexere modellen houden rekening met variatie in vlekgrootte vlekkwaliteit ruimtelijke configuratie interne dynamiek

119
Metapopulatie theorie
Aanzienlijke variatie in vlekgrootte of -kwaliteit kan aanleiding geven tot mainland-island metapopulaties of source-sink metapopulaties.

120
Metapopulatie theorie
Mainland-island model: kans op uitsterven van ‘mainland’ populatie wordt zeer klein en alle immigranten zijn ervan afkomstig.

121
Metapopulatie theorie
Mainland-island model: kans op uitsterven van ‘mainland’ populatie wordt zeer klein en alle immigranten zijn ervan afkomstig. Vergelijking: dp/dt = k.(1 - p) - e.p
 - e.p.")
122
Metapopulatie theorie
Mainland-island model: kans op uitsterven van ‘mainland’ populatie wordt zeer klein en alle immigranten zijn ervan afkomstig. Vergelijking: dp/dt = k.(1 - p) - e.p Evenwicht: p = k/(k + e)
 - e.p. Evenwicht: p = k/(k + e)")
123
Metapopulatie theorie
Source-sink model: habitat wordt ingedeeld in source (hoge kwaliteit, geboortes > sterftes) en sinks (lage kwali-teit, geboortes < sterftes).
 en sinks (lage kwali-teit, geboortes < sterftes).")
124
Metapopulatie theorie
Conclusies voor natuurbehoud

125
Metapopulatie theorie
Conclusies voor natuurbehoud Zowel nood aan bezet als onbezet habitat

126
Metapopulatie theorie
Conclusies voor natuurbehoud Zowel nood aan bezet als onbezet habitat Optimale ruimtelijke structuur is steeds een ‘compromis’

127
Metapopulatie theorie
Conclusies voor natuurbehoud Zowel nood aan bezet als onbezet habitat Optimale ruimtelijke structuur is steeds een ‘compromis’ Nood aan bescherming van source habitat

131
Metapopulatie theorie
Evolutionair-ecologisch

132
Metapopulatie theorie
Evolutionair-ecologisch Wanneer selectiedrukken verschillen tussen sources en sinks zullen individuen uit source populaties gemiddeld een hogere lokale adaptatie vertonen wegens asymme-trische gene flow.

133
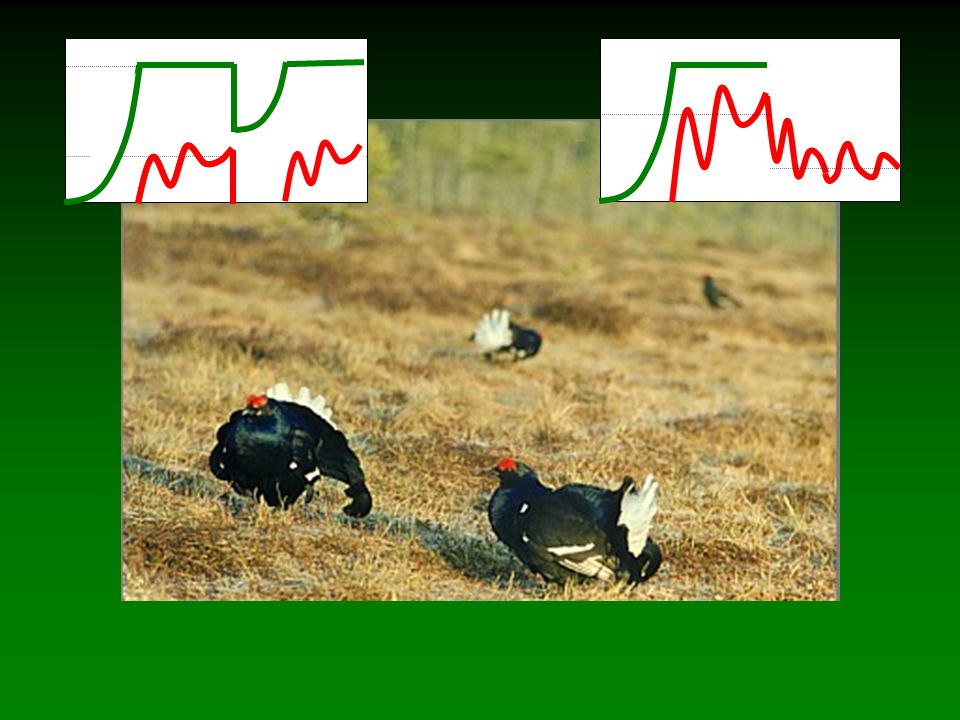
Q. pubescens Q. ilex

134
Q. pubescens bladverliezend veel voedsel Q. ilex immergroen weinig voedsel

135
Q. pubescens bladverliezend veel voedsel reproductie hoog SOURCE Q. ilex immergroen weinig voedsel reproductie laag SINK

136
voedsel Datum Q. pubescens bladverliezend veel voedsel
reproductie hoog SOURCE Q. ilex immergroen weinig voedsel reproductie laag SINK Datum voedsel

137
Corsica voedsel Datum Q. pubescens bladverliezend veel voedsel
reproductie hoog SOURCE Q. ilex immergroen weinig voedsel reproductie laag SINK Datum voedsel

138
zuid Frankrijk geen verschil in legdatum voedsel Datum Q. pubescens
bladverliezend veel voedsel reproductie hoog SOURCE Q. ilex immergroen weinig voedsel reproductie laag SINK Datum voedsel

Verwante presentaties