Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Het oog

2
PUPIL SCLERA IRIS

3
Diafragma (iris)
")
5
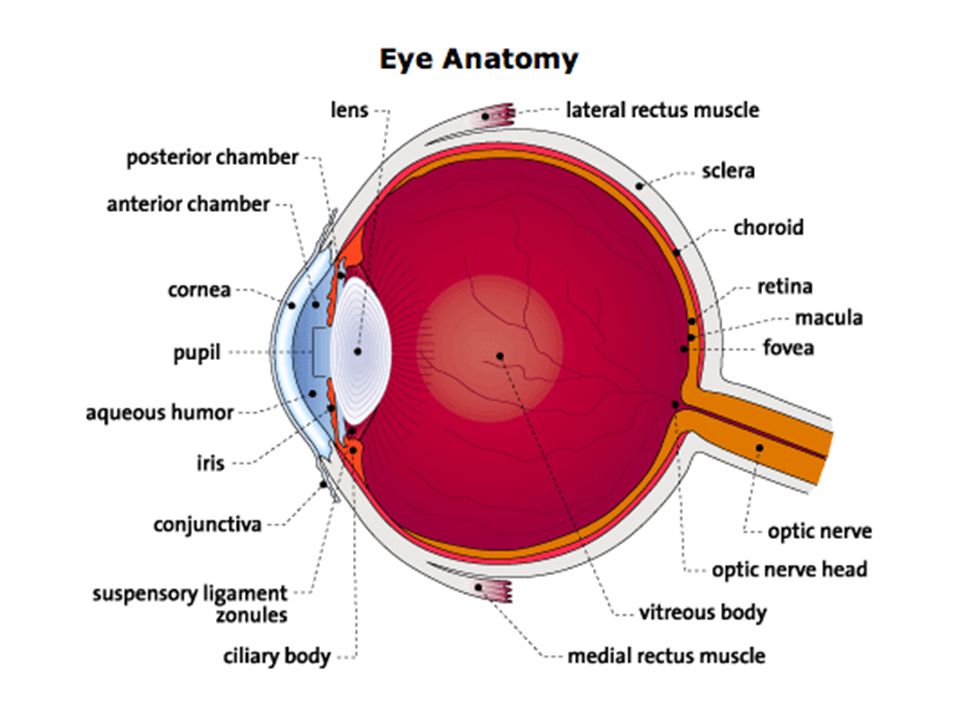
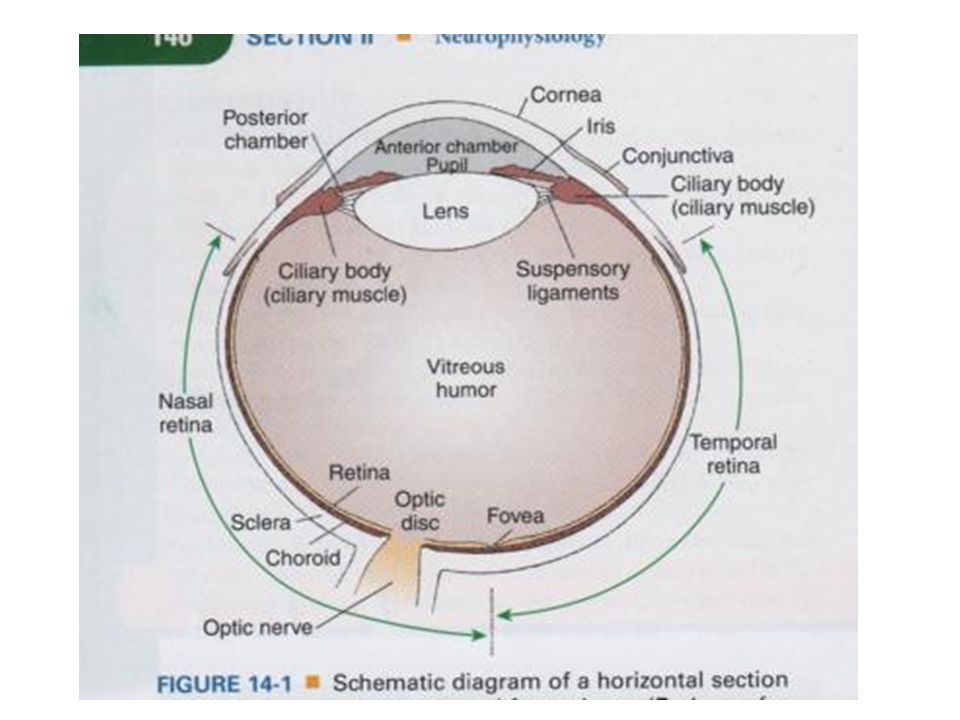
Kort overzicht anatomie van het oog
Wand oogbol: 3 lagen (oogrokken) Sclera (harde oogrok) : stevig, with ondoorzichtig vlies waaraan oogspieren zijn bevestigd (‘wit van het oog’) Gaat aan voorzijde over in de doorzichtige cornea (hoornvlies)
 Sclera (harde oogrok) : stevig, with ondoorzichtig vlies waaraan oogspieren zijn bevestigd (‘wit van het oog’) Gaat aan voorzijde over in de doorzichtige cornea (hoornvlies)")
6
Kort overzicht anatomie van het oog
Wand oogbol: 3 lagen (oogrokken) Choroidea (vaatvlies) : concentrisch met de sclera, sterk doorbloed T.h.v. overgang sclera-cornea zet het vaatvlies zich door in de iris (regenboogvlies) Iris bevat pigment Pupil kan worden vergoot/verkleind door aanpassing iris door spieren Op plaats waar choroidea in iris overgaat: verdikking van spieren: straalvormig lichaam of corpus ciliare (draagt de lens)
 Choroidea (vaatvlies) : concentrisch met de sclera, sterk doorbloed. T.h.v. overgang sclera-cornea zet het vaatvlies zich door in de iris (regenboogvlies) Iris bevat pigment. Pupil kan worden vergoot/verkleind door aanpassing iris door spieren. Op plaats waar choroidea in iris overgaat: verdikking van spieren: straalvormig lichaam of corpus ciliare (draagt de lens)")
7
Vormverandering lens : focus

8
Kort overzicht anatomie van het oog
Wand oogbol: 3 lagen (oogrokken) Retina (netvlies, binnenste oogrok): bevat de fotoreceptoren (en zenuwcellen en bloedvaten) Blinde vlek (optic disc): plaats waar oogzenuw het oog verlaat en de retina dus onderbroken is (bevat geen fotoreceptoren) Macula lutea (gele vlek) en fovea : structuur van retina waar geen zenuwuitlopers en bloedvaatjes zitten, dus enkel fotoreceptoren (zie verder)
 Retina (netvlies, binnenste oogrok): bevat de fotoreceptoren (en zenuwcellen en bloedvaten) Blinde vlek (optic disc): plaats waar oogzenuw het oog verlaat en de retina dus onderbroken is (bevat geen fotoreceptoren) Macula lutea (gele vlek) en fovea : structuur van retina waar geen zenuwuitlopers en bloedvaatjes zitten, dus enkel fotoreceptoren (zie verder)")
10
Kort overzicht anatomie van het oog
Glasachtig lichaam (corpus vitreum): glasheldere, geleiachtige vloeistof Kamervocht: waterheldere vloeistof tussen cornea en lens Voorste : tussen cornea en iris Achterste: tussen iris, lens, corpus ciliare en ligamenten van lens
: glasheldere, geleiachtige vloeistof. Kamervocht: waterheldere vloeistof tussen cornea en lens. Voorste : tussen cornea en iris. Achterste: tussen iris, lens, corpus ciliare en ligamenten van lens.")
12
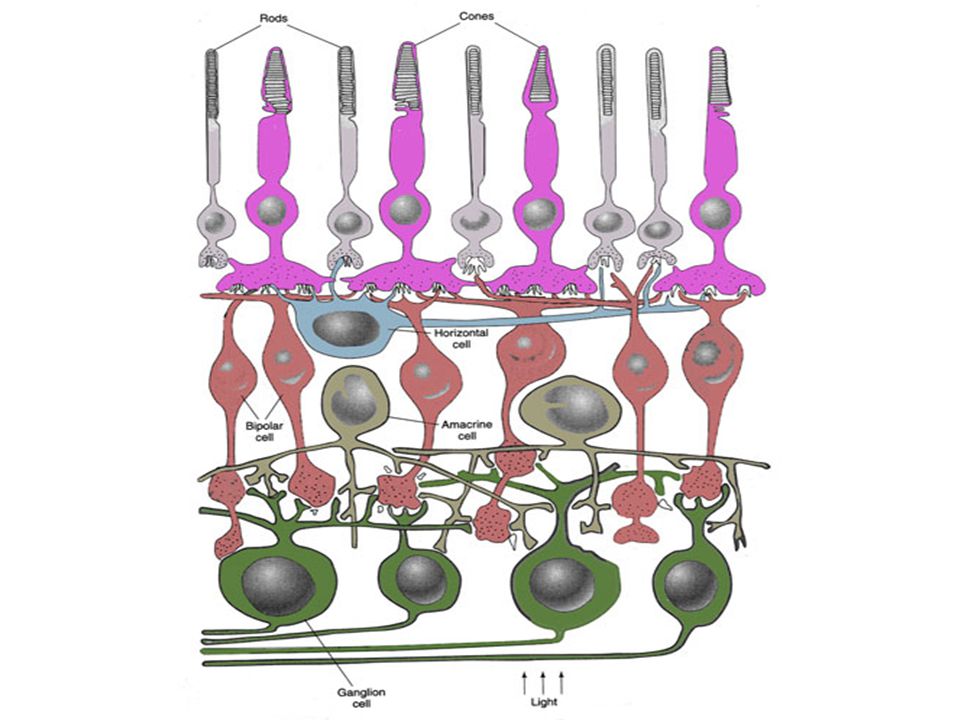
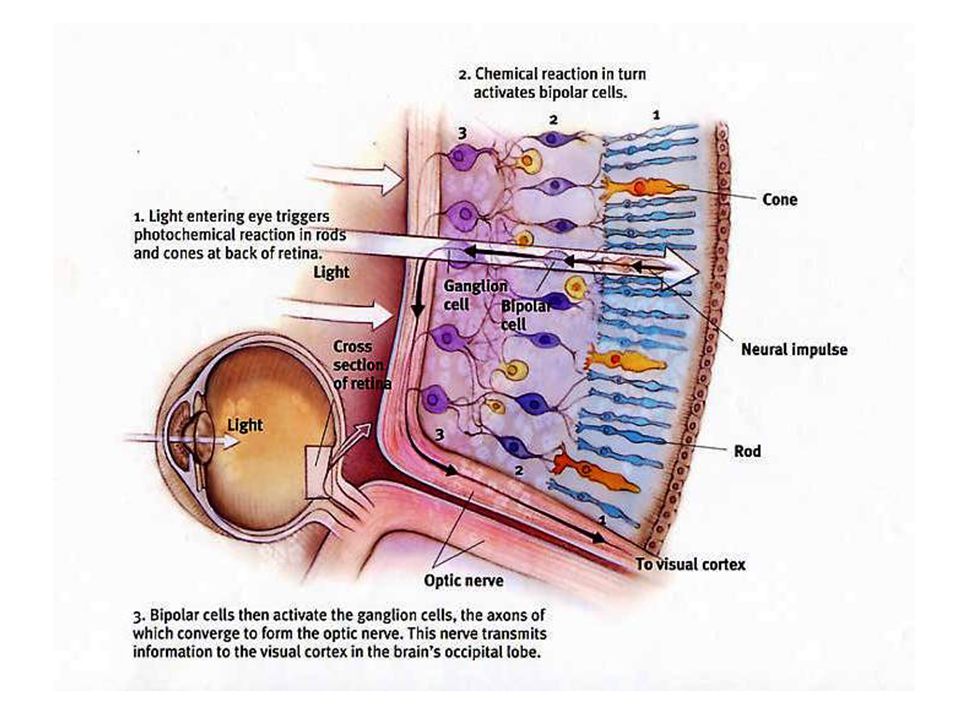
Het netvlies (retina) : celtypes
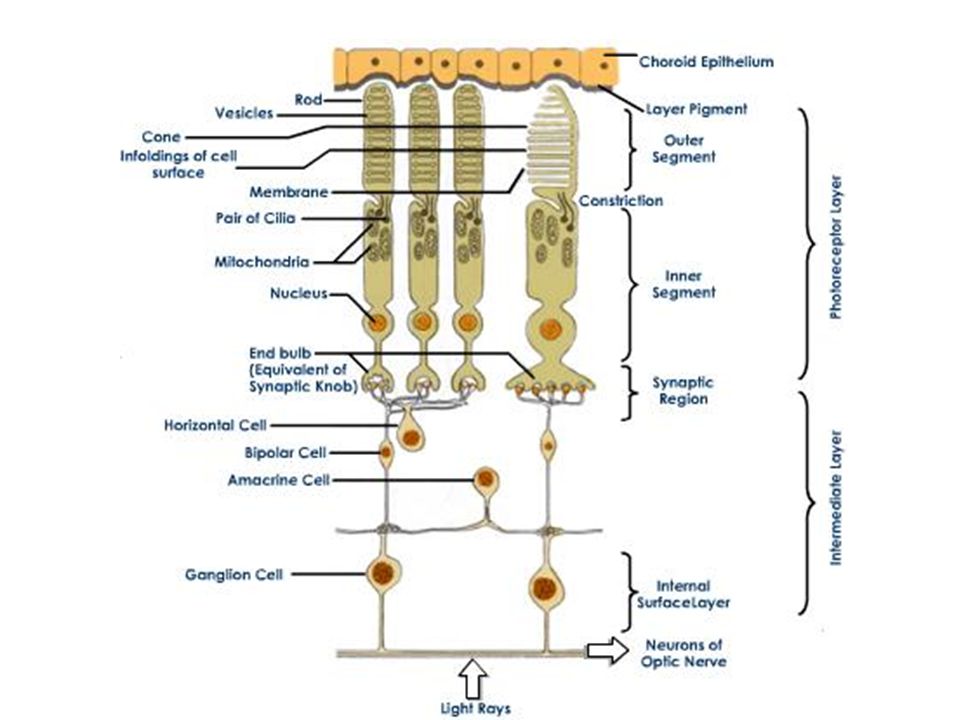
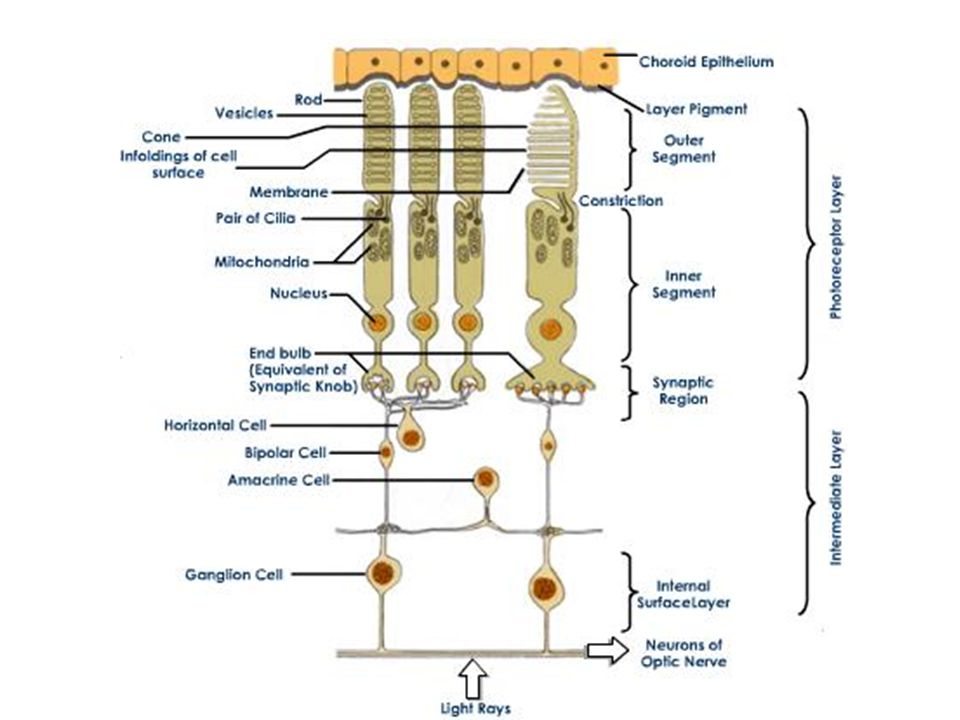
Retina afkomstig van neuroectoderm: oog is extensie van hersenen 5 belangrijke celtypes: Fotoreceptorcellen (kegeltjes en staafjes) Bipolaire cellen Horizontale cellen Amacrine cellen Ganglion cellen interneuroncellen
 Bipolaire cellen. Horizontale cellen. Amacrine cellen. Ganglion cellen. interneuroncellen.")
14
Het netvlies (retina) : celtypes
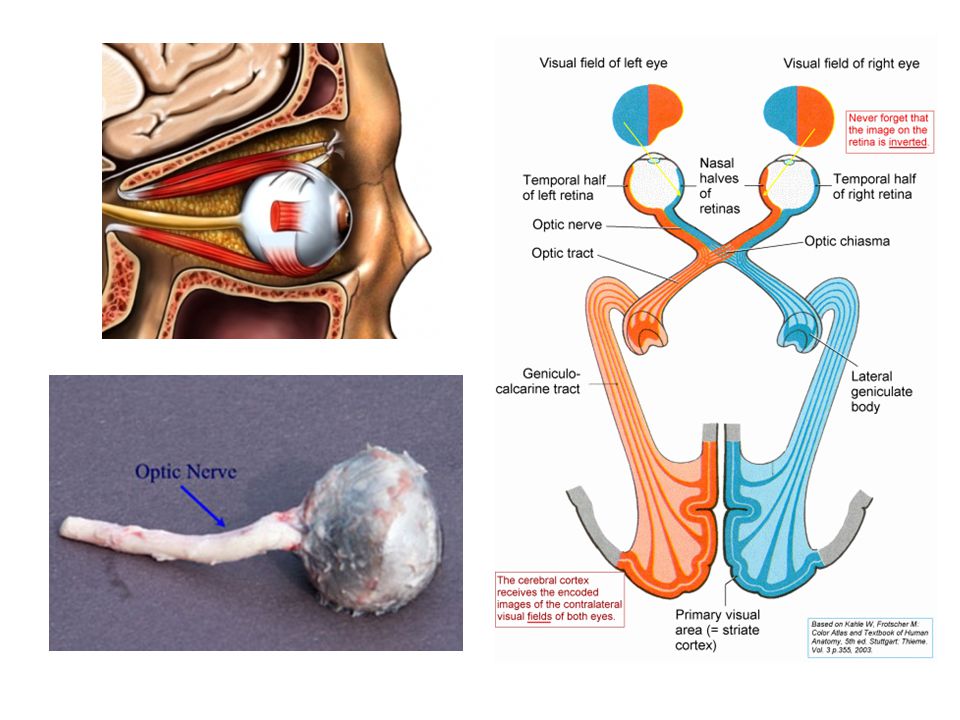
Fotoreceptorcellen maken synaptische verbinding met bipolaire cellen Bipolaire cellen maken contact met ganglioncellen Ganglioncellen komen samen thv blinde vlek en sturen via optische zenuwbanen signalen naar hersenen

15
Het netvlies (retina) : celtypes
De horizontale en amacriene cellen mediëren laterale interacties tussen A) fotoreceptoren en bipolaire cellen B) bipolaire cellen en ganglioncellen
 fotoreceptoren en bipolaire cellen. B) bipolaire cellen en ganglioncellen.")
16
Het netvlies (retina) : celtypes
Retinale ganglion cellen bevinden zich aan binnenkant retina (bij glasachtig lichaam) Fotoreceptoren bevinden zich aan buitenkant retina (bij choroidea) Lichtdistortie !!
 Fotoreceptoren bevinden zich aan buitenkant retina (bij choroidea) Lichtdistortie !!")
18
Het netvlies (retina) : celtypes
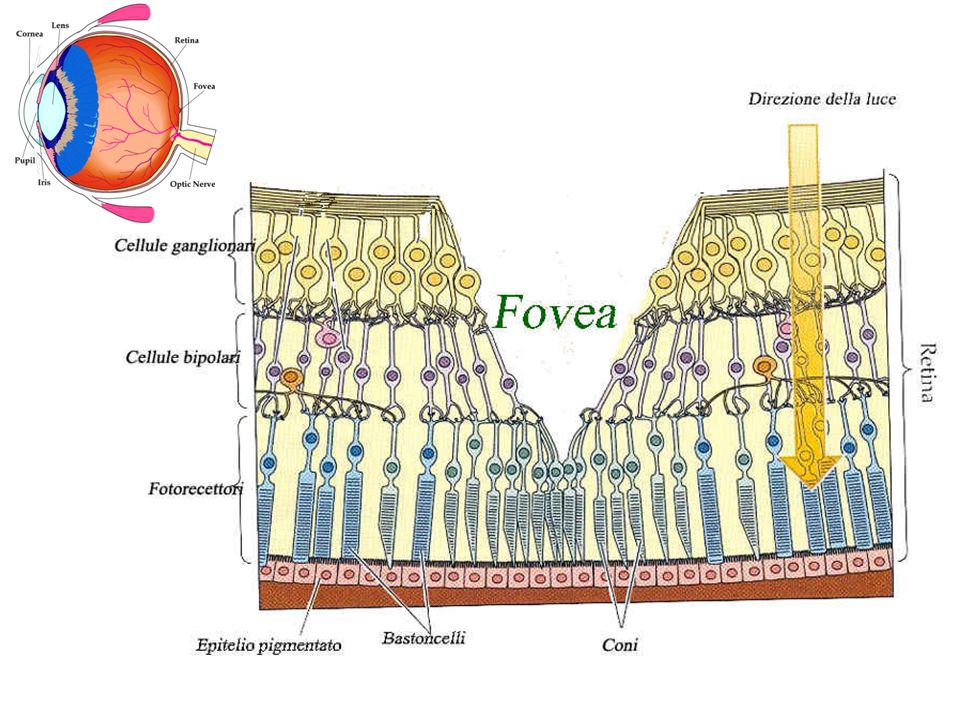
Oplossing probleem van lichtdistortie: fovea Geen ganglioncellen en bipolaire cellen Enkel fotoreceptorcellen: staafjes en kegeltjes Geen obstructie in toegang licht tot fotoreceptoren Zone met hoogste visuele activiteit

20
Retina en pigment Dagdieren: donker pigment in epitheel tussen fotoreceptoren en vaatvlies Lichtabsorptie, geen reflectie Nachtdieren: reflecterend pigment (tapetum) Optimaal gebruik van licht (maar minder scherp)
 Optimaal gebruik van licht (maar minder scherp)")
22
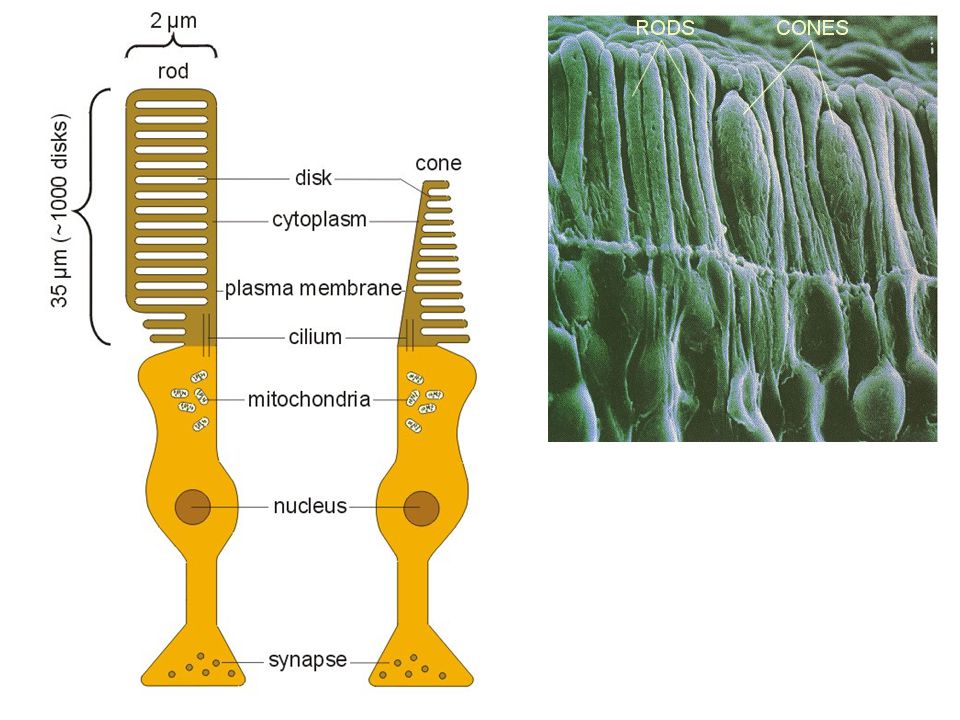
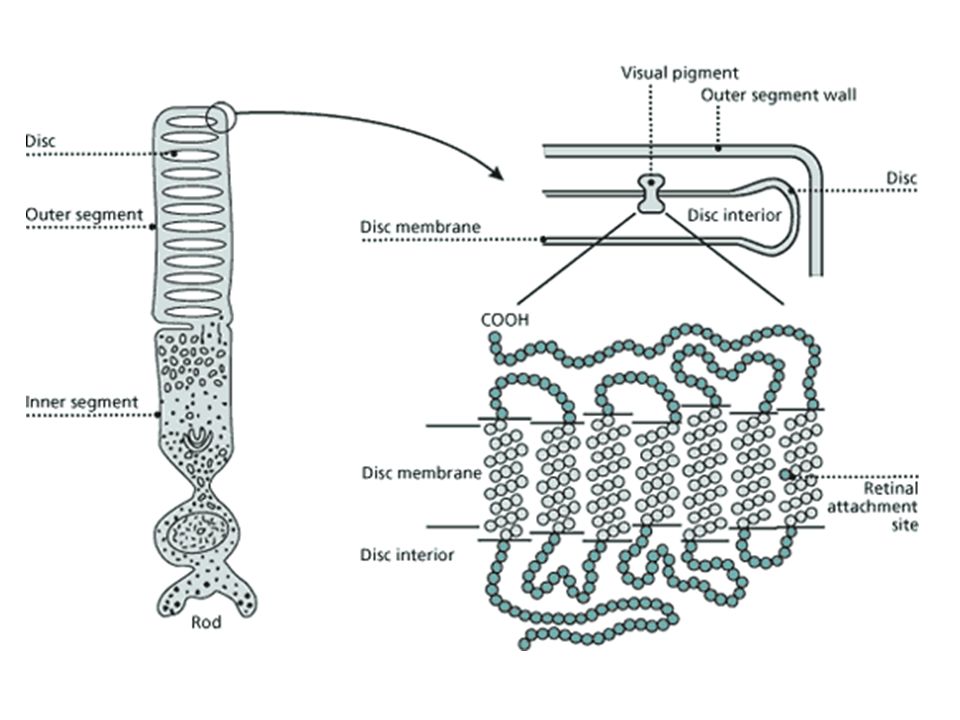
Fotoreceptorcellen Staafjes (rods) en kegeltjes (cones): neuronale cellen Driedelig: Synaptisch einde: synaps met bipolaire cellen Binnenste segment: kern, mitochondria, andere cytoplasmatische structuren Buitenste segment: membraandisks met actieve lichtgevoelige moleculaire complexen, die licht omzetten in neuronale electrische signalen (en continue vernieuwing discs) Cilium verbindt binnenste en buitenste segment
 Cilium verbindt binnenste en buitenste segment.")
24
Fotoreceptorcellen Staafjes (rods): nacht-zicht
zeer gevoelig aan licht Meerder staafjes voeden synaptische input naar 1 bipolaire cel Geen scherp beeld, geen kleurzicht Kegeltjes (cones): dag-zicht Minder lichtgevoelig 1 (of slechts enkele) kegeltjes voeden 1 bipolaire cel Scherp beeld Kleurzicht (verschillende fotopigmenten, actief mbt verschillende golflengtes licht)
: dag-zicht. Minder lichtgevoelig. 1 (of slechts enkele) kegeltjes voeden 1 bipolaire cel. Scherp beeld. Kleurzicht (verschillende fotopigmenten, actief mbt verschillende golflengtes licht)")
27
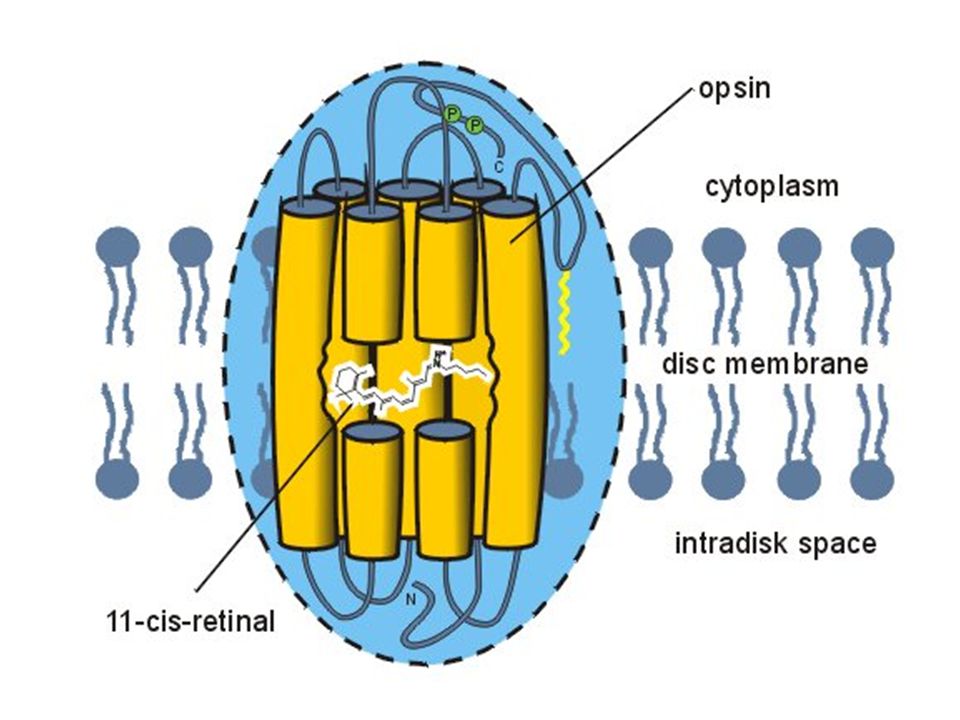
Biochemie van het oog Fotochemisch actief complex = rhodopsine
Rhodopsine = combinatie van opsine (een proteïne) en 11-cis-retinal hv 11-cis-retinal all-trans-retinal Isomerisatie
 en 11-cis-retinal. hv. 11-cis-retinal all-trans-retinal. Isomerisatie.")
28
Biochemie van het oog Retinals gerelateerd aan retinolen (vitamine A)
Geen de novo synthese mogelijk Vorming door afbraak van carotenoïden, zoals β-caroteen

29
Biochemie van het oog Opsine = intrinsiek membraanproteïne uit disk membranen (› 95% van eiwitfractie) C-terminaal deel in cytoplasma; N-terminaal deel in ruimte in microvesiculae (disc) 7 hydrofobe transmembraan α-helices
 7 hydrofobe transmembraan α-helices.")
32
Rhodopsine Retinal gebonden aan lysyl-NH2 groep via aldimine (R–CH=N–R') of geprotoneerde Schiff base binding
 of geprotoneerde Schiff base binding.")
33
Fotoisomerisatie van rhodopsine
Isomerisatie 11-cis-retinal tot all-trans-retinal oiv licht leidt to conformatieverandering rhodopsine

34
Staafjes vs kegeltjes Rhodopsine is fotoactieve component van staafjes

35
Staafjes vs kegeltjes Rhodopsine is fotoactieve component van staafjes

36
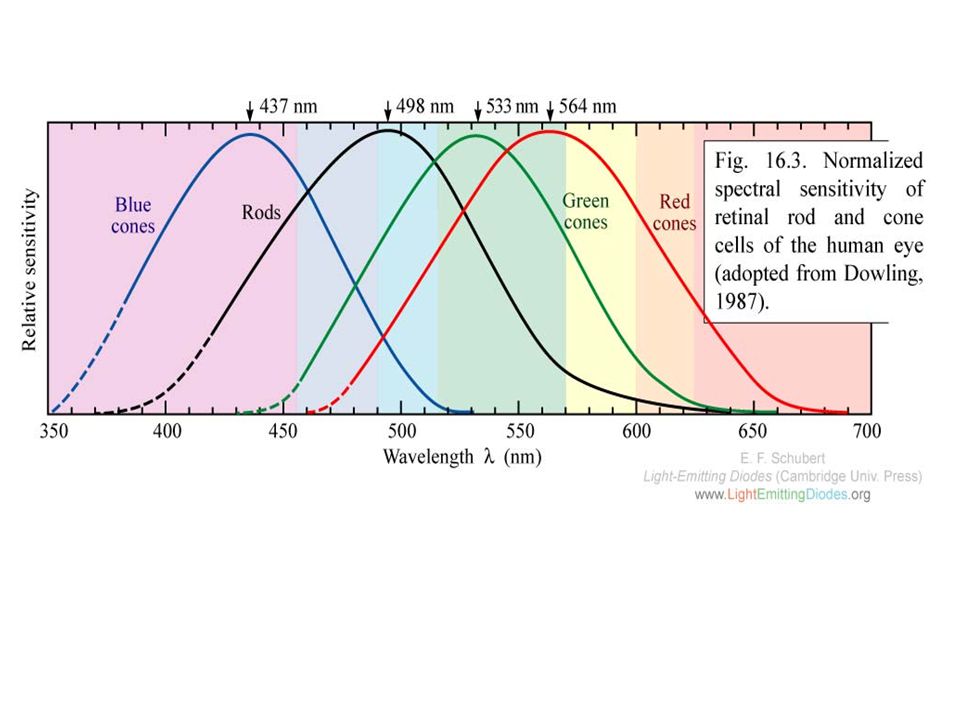
Staafjes vs kegeltjes Kegeltjes : 3 verschillende soorten
Bevatten allen 11-cis-retinal gebonden aan proteïnen gelijkend op opsine: verschillende spectra = iodopsines (of photopsines)
")
37
Fotochemische transformatie van rhodopsine

38
Fotochemische transformatie van rhodopsine
Rhodopsine transformeert tot metarhodopsine II via reeks metastabiele moleculen Transformatie gekoppeld met verschuiving ladingen van Schiff’s base over 11-cis retinal Ladingsverschuiving gekoppeld aan geometrische veranderingen opsine (rhodopsine)
")
39
Verschuiving van de positieve lading over de ring door excitatie van de geprotoneerde Schiffse base

40
Fotochemische conversie van rhodopsine naar bathorhodopsine

41
Fotochemische transformatie van rhodopsine
Vrijkomen van all-trans retinal van opsine duurt lang: dus geen essentiële stap in zicht Stap belangrijk in regeneratie van de actieve vorm van rhodopsine (vorming opsine en all-trans retinal) All-trans retinal wordt omgezet tot 11-cis retinal via een retinyl-ester als intermediair
 All-trans retinal wordt omgezet tot 11-cis retinal via een retinyl-ester als intermediair.")
42
Regeneratie van 11-cis retinal

43
Regeneratie van 11-cis retinal
Reductie van all-trans-retinal tot all-trans-retinol via NADH All-trans-retinol verlaat netvlies en wordt opgenomen door pigmentepitheel Verestering door acyltransferase: transfer van vetzuur van phosphatidylcholine aan all-trans-retinol: vorming van een all-trans-retinylester

44
Regeneratie van 11-cis retinal
All-trans retinylester hydrolyseert tot 11-cis retinol en het vrije vetzuur (isomerisatiestap van all-trans retinol tot 11-cis retinol) Regeneratie van 11-cis retinal uit 11-cis retinol door oxidatie dmv NAD+ Migratie van 11-cis retinal van pigmentepitheel naar netvlies, binding aan opsine = rhodopsine
 Regeneratie van 11-cis retinal uit 11-cis retinol door oxidatie dmv NAD+ Migratie van 11-cis retinal van pigmentepitheel naar netvlies, binding aan opsine = rhodopsine.")
45
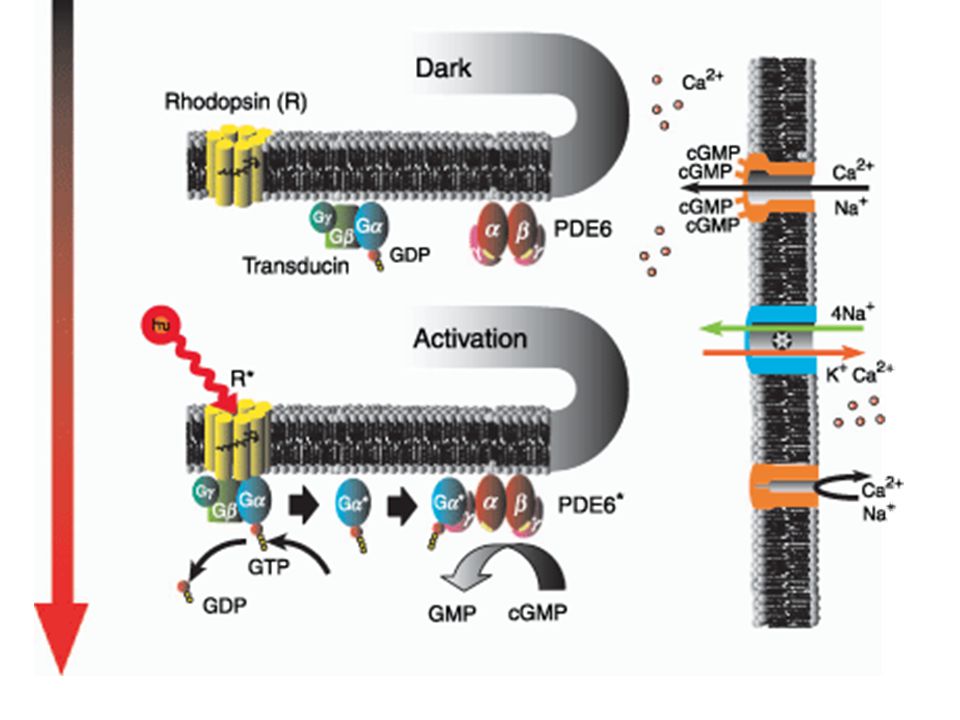
Elektrofysiologische respons van fotoreceptorcellen

46
Elektrofysiologische respons van fotoreceptorcellen
Respons op licht = verandering in permeabiliteit van cytoplasmatische membraan voor kationen Cytoplasmamembraan op binnenste segment bevat Na+-K+ pomp Na+-K+ pomp gebruikt ATP om Na+ naar buiten en K+ naar binnen te pompen

47
Elektrofysiologische respons van fotoreceptorcellen
K+ kan vrij uit cel diffunderen in binnenste segment Na+ kan niet vrij diffunderen naar cytoplasma Na+ migreert naar buitenste segment en treedt binnen via een kation selectief kanaal Migreert dan naar binnenste segment en de cyclus herbegint

48
Elektrofysiologische respons van fotoreceptorcellen

49
Elektrofysiologische respons van fotoreceptorcellen
Licht veroorzaakt sluiting van kation selectieve kanalen in buitenste segment Inwaardse Na+-stroom beperkt Potentiaalverschil over cytoplasmatische membraan vergroot = Hyperpolarisatie

51
Hyperpolarisatie en synaps
Productie neurotransmitters naar bipolaire cellen Signaal van bipolaire cel naar ganglioncel Actiepotentiaal van ganglioncellen sturen signaal naar hersenen

53
Rhodopsine activatie op membraan microvesiculae (disc membranes)
Hyperpolarisatie door sluiting kation kanalen op cytoplasmatische membraan

54
Rhodopsine activatie op membraan microvesiculae (disc membranes)
?? Rhodopsine activatie op membraan microvesiculae (disc membranes) ?? Hyperpolarisatie door sluiting kation kanalen op cytoplasmatische membraan
 Hyperpolarisatie door sluiting kation kanalen op cytoplasmatische membraan.")
55
cGMP (3’-5’ cyclisch GMP)
cGMP veroorzaakt een stijging in membraan-permeabiliteit voor Na+, interactie met Na+ kanalen op buitenste segment cGMP diffundeert tussen membraan van microvesiculae en cytoplamatische membraan

56
Phosphodiesterase Omzetting cGMP naar 5’ GMP op membraan van microvesiculae (disc membrane) + H H+ cGMP 5’ GMP

57
Phosphodiesterase Omzetting cGMP naar 5’ GMP op membraan van microvesiculae (disc membrane)
")
58
Activatie rhodopsine, vorming metarhodopsine II
Licht Activatie rhodopsine, vorming metarhodopsine II Activatie phosphodiesterase Afbraak cGMP Sluiting Na+ kanaal in buitenste segment Potentiaalverschil, synaps

59
Phosphodiesterase Omzetting cGMP naar 5’ GMP op membraan van microvesiculae (disc membrane) Activatie door licht heeft GTP nodig en een ander membraanproteïne : transducine Transducine is een G-protëine

60
G-proteïne Actieve component

61
cGMP Rhodopsin 5’ GMP

62
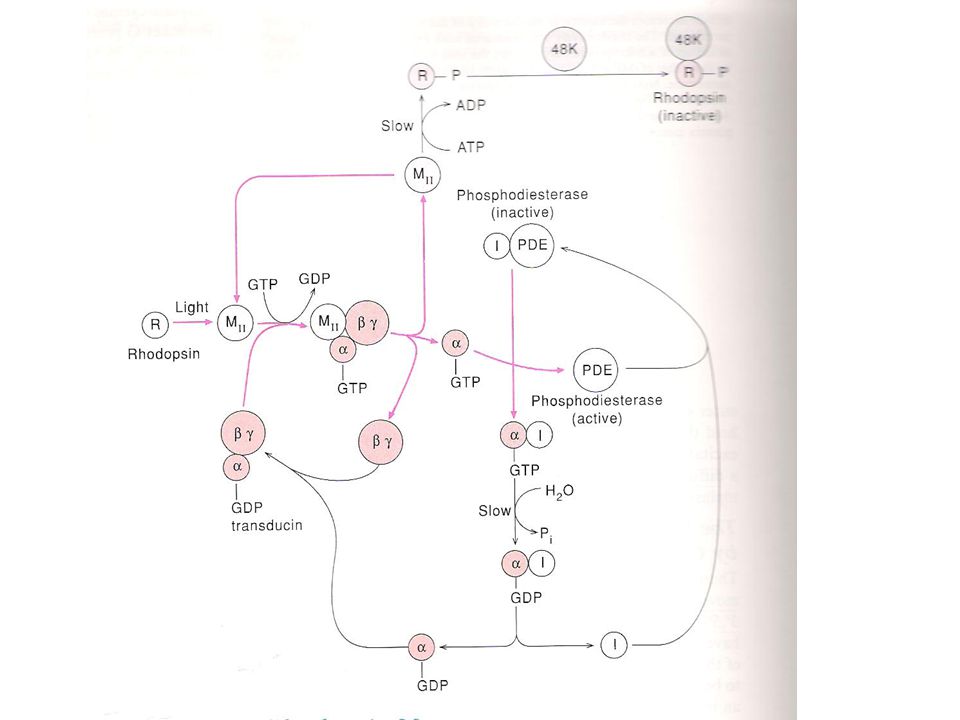
Werking transducine Transducine bestaat uit α,β en γ subeenheden
Metarhodopsine interageert met transducine, met als gevolg verplaatsen van GDP door GTP op α-subeenheid Afscheiding van α subeenheid van β en γ subeenheden Binden van α subeenheid aan inhibitorische component van phosphodiesterase Activatie van phosphodiesterase

63
Amplificatie van lichtsignaal
Eén molecule metarhodopsine activeert ± 500 transducine moleculen Eén molecule phosphodiesterase hydrolyseert meerdere moleculen cGMP (meerdere staafjes voeden een bipolaire cel)
")
65
Stopzetten signaal Hydrolyse van aan α subeenheid gebonden GTP naar GDP, gevolgd door vrijkomen van inhibitorische component en binding aan phosphodiesterase (inactivatie PDE) Verval van metarhodopsine II naar rhodopsine(via enzyme (48K) arrestine) Regeneratie van cGMP door guanylate cyclase (via omzetting GTP naar cGMP)
 Verval van metarhodopsine II naar rhodopsine(via enzyme (48K) arrestine) Regeneratie van cGMP door guanylate cyclase (via omzetting GTP naar cGMP)")
Verwante presentaties









 Zintuig = orgaan dat reageert op prikkels uit het milieu>")











