Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
5. Ontwikkelingen in de twintigste eeuw
Van artificieel klasseren naar fylogenetische reconstructie Historische ontwikkeling van de systematiek 5. Ontwikkelingen in de twintigste eeuw karyosystematiek chemotaxonomie serologische systematiek moleculaire systematiek experimentele systematiek numerieke taxonomie cladisme Ontwikkelingen van de twintigste eeuw In de twintigste eeuw groeit de polemiek of classificaties fenetisch dan wel fylogenetisch moeten zijn. De fenetici hielden voor dat fossiele gegevens voor altijd zo fragmentair zouden blijven dat een nauwkeurige reconstructie van de evolutie voor de meeste organismen toch onhaalbaar is. Een geperfectioneerde fenetische systematiek werd dan geacht best gebaseerd te zijn op een zeer groot aantal kenmerken; zulke fenetische systemen zouden vanzelf in hoge mate congruent zijn met de afstammingsgeschiedenis. Voor het verwerken van grote hoeveelheden gegevens zijn de opvattingen en methoden van de numerieke taxonomie in het bijzonder geschikt. Ze kwam tot ontwikkeling in de systematische microbiologie, waar grote en consistente sets van gegevens werden bekomen bij gekweekte bacteriën en andere micro-organismen. Daarnaast is vanaf de jaren veertig de biosystematiek of experimentele systematiek tot ontwikkeling gekomen. Deze `New Systematics’ maakt gebruik van genetische en ecologische experimenten en karyologische waarnemingen. Vanaf dezelfde periode zullen ook chemische gegevens het aantal kenmerken voor systematische evidentie sterk verbreden. In de plantkunde worden secundaire plantenstoffen het voorwerp van chemotaxonomie. In de dierkunde heeft de vergelijkende studie van eiwitten meer succes gekend (serologische systematiek). In de jaren zestig komt de cladistiek of het cladisme tot ontwikkeling. Deze denkrichting postuleert dat niet zozeer veel kenmerken moeten bestudeerd worden, als wel de evolutie van de kenmerken moet geanalyseerd worden. Het gemeenschappelijk bezit van afgeleide kenmerktoestanden is dan sluitende bewijs voor gemeenschappelijke afstamming. Deze “beweging” kwam tot stand in de entomologie en postuleert dus dat de genealogische vertakkingspatronen van de evolutie kunnen gereconstrueerd worden zonder fossielen. In de jaren tachtig zal de cladistiek definitief zijn plaats verwerven in de plantkunde, waar de methodiek lang zijn sterkste opponenten vond. Tenslotte laten ook de spectaculaire ontwikkelingen van de moleculaire biologie en van de biotechnologie moeder systematiek niet onberoerd; ze verschaffen immers het middel taxa te vergelijken via de directe vergelijking van hun genoom. Voor de zaadplanten heeft sequentiebepaling van vooral delen van chloroplast‑DNA zoveel resultaten opgeleverd dat “fylogenie” welhaast synoniem is geworden van “fylogenie afgeleid uit macromoleculaire gegevens.” De cladistische algoritmen bewezen de meest geëigende verwerkingsmethode te zijn voor de gegevenssets van deze richting die vaak kortweg aangeduid wordt als “moleculaire systematiek.”
. In de jaren zestig komt de cladistiek of het cladisme tot ontwikkeling. Deze denkrichting postuleert dat niet zozeer veel kenmerken moeten bestudeerd worden, als wel de evolutie van de kenmerken moet geanalyseerd worden. Het gemeenschappelijk bezit van afgeleide kenmerktoestanden is dan sluitende bewijs voor gemeenschappelijke afstamming. Deze beweging kwam tot stand in de entomologie en postuleert dus dat de genealogische vertakkingspatronen van de evolutie kunnen gereconstrueerd worden zonder fossielen. In de jaren tachtig zal de cladistiek definitief zijn plaats verwerven in de plantkunde, waar de methodiek lang zijn sterkste opponenten vond. Tenslotte laten ook de spectaculaire ontwikkelingen van de moleculaire biologie en van de biotechnologie moeder systematiek niet onberoerd; ze verschaffen immers het middel taxa te vergelijken via de directe vergelijking van hun genoom. Voor de zaadplanten heeft sequentiebepaling van vooral delen van chloroplast‑DNA zoveel resultaten opgeleverd dat fylogenie welhaast synoniem is geworden van fylogenie afgeleid uit macromoleculaire gegevens. De cladistische algoritmen bewezen de meest geëigende verwerkingsmethode te zijn voor de gegevenssets van deze richting die vaak kortweg aangeduid wordt als moleculaire systematiek.")
2
Ontwikkeling van experimentele richtingen in de biologie
Begin twintigste eeuw Ontwikkeling van experimentele richtingen in de biologie Systematiek ‘dormant’ Kopieer materiaal uit lezing lawalree (dumortiera)
")
3
1665 Robert Hooke ontdekking van de cel
1838 M.J. Scheiden celtheorie 1882 W. Flemming ontdekking van structuren die overlangs splitsen tijdens mitose (helften gaan naar dochtercellen) Waldeyer benaming 'chromosoom‘ >>>>> beschrijvende cytologie
 Waldeyer benaming chromosoom‘ >>>>> beschrijvende cytologie.")
4
Menselijke geslachtschromosomen
Nature (2003) 423 :
 423 :")
5
De Vries herontdekking mendelisme
>>>> experimentele cytologie & cytogenetica

6
Julian Huxley (1940) The New Systematics
Julian Huxley came from the distinguished Huxley family. His brother was the writer Aldous Huxley, and his half-brother a fellow biologist and Nobel laureate, Andrew Huxley; his father was writer and editor Leonard Huxley; and his paternal grandfather was Thomas Henry Huxley, a friend and supporter of Charles Darwin and protagonist of evolution. His maternal grandfather was the academic Tom Arnold, great-uncle poet Matthew Arnold and great-grandfather Thomas Arnold of Rugby School.

8
Cytologische waarnemingen
Aantal Vorm Relatieve lengte Techniek: Squash-preparaat (mitose of meiose)
")
9
Meiose in d, andere preparaten mitose
Bij hoge chromosoomgetallen is het voordelig meioses te bestuderen

10
Enkele voorbeelden van chromosoomgetallen (2n) bij planten
Arabidopsis thaliana Brassicac. 2 Haplopappus gracilis Asterac. 2 of 4 (rassen!) Crepis neglecta Asterac. 8 Crepis kotschyana Asterac. 8 Crepis fuliginosa Asterac. 22 Sagittaria, alle spp. Alismatac. 22 Pinus, alle spp. Gymnospermae 24 Caltha palustris Ranunculac. 32 of 56 Crepis leontodontoides Asterac. 38 Crepis kashmirica Asterac. 100 Poa litorosa Poac Voanicola Arecac. 450 Ophioglossum reticulatum varens c. 1400 sommige genera variabel, andere constant 2. Grootteorde vergelijkbaar met dieren (mens 2n = 46) 3. Record bloemplanten: Voanicola (endemische palm Madagascar; bepaling dateert van ca. 1990) 4. varens hebben zeer hoge ‘secundaire’ chromosoomgetallen
 Crepis neglecta Asterac. 8. Crepis kotschyana Asterac. 8. Crepis fuliginosa Asterac. 22. Sagittaria, alle spp. Alismatac. 22. Pinus, alle spp. Gymnospermae 24. Caltha palustris Ranunculac. 32 of 56. Crepis leontodontoides Asterac. 38. Crepis kashmirica Asterac Poa litorosa Poac Voanicola Arecac Ophioglossum reticulatum varens c sommige genera variabel, andere constant. 2. Grootteorde vergelijkbaar met dieren (mens 2n = 46) 3. Record bloemplanten: Voanicola (endemische palm Madagascar; bepaling dateert van ca. 1990) 4. varens hebben zeer hoge ‘secundaire’ chromosoomgetallen.")
11
Brachycome dichromosomatica 2n = 4
Brachycome (Asteraceae) Ophioglossum (varens) Ophioglossum reticulatum 2n = c. 1400
 Ophioglossum (varens) Ophioglossum reticulatum. 2n = c")
12
Enkele voorbeelden van chromosoomgetallen (2n) bij planten
Arabidopsis thaliana Brassicac. 2 Haplopappus gracilis Asterac. 2 of 4 (rassen!) Crepis neglecta Asterac. 8 Crepis kotschyana Asterac. 8 Crepis fuliginosa Asterac. 22 Sagittaria, alle spp. Alismatac. 22 Pinus, alle spp. Gymnospermae 24 Caltha palustris Ranunculac. 32 of 56 Crepis leontodontoides Asterac. 38 Crepis kashmirica Asterac. 100 Poa litorosa Poac Voanicola Arecac. 450 Ophioglossum reticulatum varens 1260 sommige genera variabel, andere constant 2. Grootteorde vergelijkbaar met dieren (mens 2n = 46) 3. Record bloemplanten: Voanicola (endemische palm Madagascar; bepaling dateert van ca. 1990 4. varens hebben zeer hoge ‘secundaire’ chromosoomgetallen
 Crepis neglecta Asterac. 8. Crepis kotschyana Asterac. 8. Crepis fuliginosa Asterac. 22. Sagittaria, alle spp. Alismatac. 22. Pinus, alle spp. Gymnospermae 24. Caltha palustris Ranunculac. 32 of 56. Crepis leontodontoides Asterac. 38. Crepis kashmirica Asterac Poa litorosa Poac Voanicola Arecac Ophioglossum reticulatum varens sommige genera variabel, andere constant. 2. Grootteorde vergelijkbaar met dieren (mens 2n = 46) 3. Record bloemplanten: Voanicola (endemische palm Madagascar; bepaling dateert van ca varens hebben zeer hoge ‘secundaire’ chromosoomgetallen.")
13
Enkele voorbeelden van chromosoomgetallen (2n) bij planten
Arabidopsis thaliana Brassicac. 2 Haplopappus gracilis Asterac. 2 of 4 (rassen!) Crepis neglecta Asterac. 8 Crepis kotschyana Asterac. 8 Crepis fuliginosa Asterac. 22 Sagittaria, alle spp. Alismatac. 22 Pinus, alle spp. Gymnospermae 24 Caltha palustris Ranunculac. 32 of 56 Crepis leontodontoides Asterac. 38 Crepis kashmirica Asterac. 100 Poa litorosa Poac Voanicola Arecac. 450 Ophioglossum reticulatum varens 1260 sommige genera variabel, andere constant 2. Grootteorde vergelijkbaar met dieren (mens 2n = 46) 3. Record bloemplanten: Voanicola (endemische palm Madagascar; bepaling dateert van ca. 1990 4. varens hebben zeer hoge ‘secundaire’ chromosoomgetallen
 Crepis neglecta Asterac. 8. Crepis kotschyana Asterac. 8. Crepis fuliginosa Asterac. 22. Sagittaria, alle spp. Alismatac. 22. Pinus, alle spp. Gymnospermae 24. Caltha palustris Ranunculac. 32 of 56. Crepis leontodontoides Asterac. 38. Crepis kashmirica Asterac Poa litorosa Poac Voanicola Arecac Ophioglossum reticulatum varens sommige genera variabel, andere constant. 2. Grootteorde vergelijkbaar met dieren (mens 2n = 46) 3. Record bloemplanten: Voanicola (endemische palm Madagascar; bepaling dateert van ca varens hebben zeer hoge ‘secundaire’ chromosoomgetallen.")
14
Vergelijkende interpretatie
van deze cytologische variatie: cytotaxonomie karyosystematiek

15
2n = 44 n = haploïd getal x = 11 (basisgetal) (auto)polyploïdie
chromosoomgetal 2n = 22 n = haploïd getal x = 11 (basisgetal) 2n = 44 n = haploïd getal x = 11 (basisgetal) (auto)polyploïdie Xxxxx toevoegen chromosoompreparaat) Tel 11, 22, 20
 2n = 44. n = haploïd getal. x = 11 (basisgetal) (auto)polyploïdie. Xxxxx toevoegen chromosoompreparaat) Tel 11, 22, 20.")
16
Sinds 1958: Index of Plant Chromosome Numbers

17
Nl 28-118 (1966) Be 16-118 (1984) Cardamine pratensis
 Be (1984) Cardamine pratensis")
18
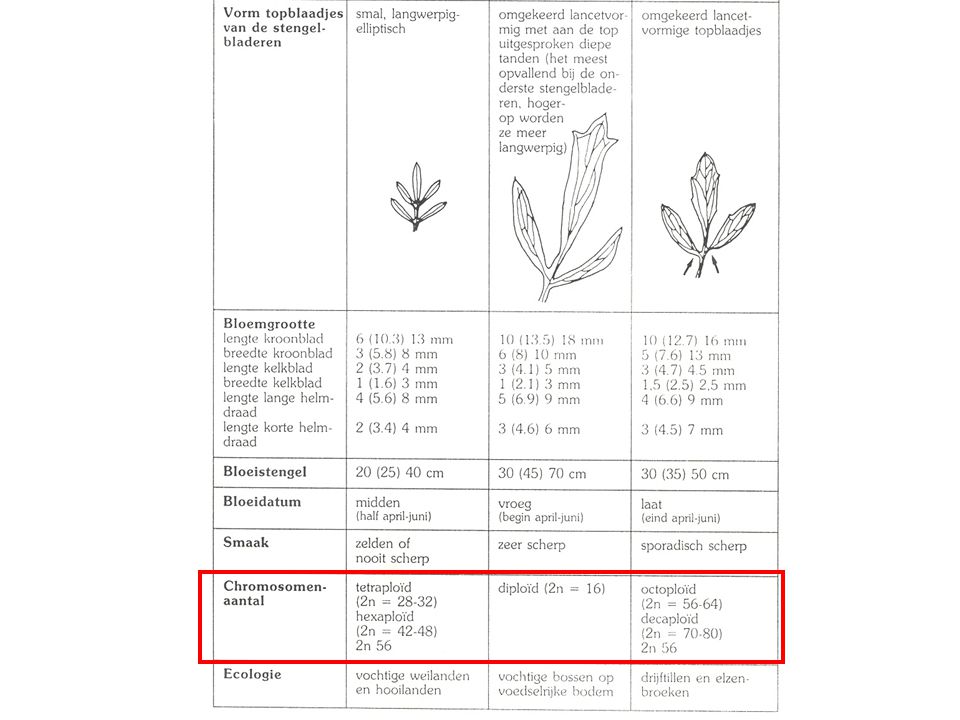
? Cardamine pratensis ploïdie 2n di- 16 tetra- 28, 30, 31, 32
hybridogeen 34, 36, 38, 39 hexa- 42, 44, 45, 46, 48 hybridogeen 52, 53, 54 octo- 56, 58, 59, 60, 61, 62, 64 hybridogeen 65, 66, 67, 68 deca- 70, 72, 73, 74, 75, 76, 78, 80, 84 ? ca. 118 ?

19
POLYPLOÏDIE Cardamine pratensis ploïdie 2n di- 16
tetra- 28, 30, 31, 32 34, 36, 38, 39 hexa- 42, 44, 45, 46, 48 52, 53, 54 octo- 56, 58, 59, 60, 61, 62, 64 65, 66, 67, 68 deca- 70, 72, 73, 74, 75, 76, 78, 80, 84 ? ca. 118 POLYPLOÏDIE

20
ANEUPLOÏDIE (polyploïdisatie + hybridisatie) Cardamine pratensis
ploïdie 2n di- 16 tetra- 28, 30, 31, 32 hybridogeen 34, 36, 38, 39 hexa- 42, 44, 45, 46, 48 hybridogeen 52, 53, 54 octo- 56, 58, 59, 60, 61, 62, 64 hybridogeen 65, 66, 67, 68 deca- 70, 72, 73, 74, 75, 76, 78, 80, 84 ? ca. 118 ANEUPLOÏDIE (polyploïdisatie + hybridisatie)
")
21
Verklaring van de onregelmatigheden:
Bastaardering tussen ploïdie-niveaus Fragmentatie of fusie van chromosomen Dysploïde translocaties B-chromosomen

24
Spartina maritima 2n = 56 (n = 28) Europa Spartina alterniflora 2n = 70 (n=35) N-Amerika
 Europa Spartina alterniflora 2n = 70 (n=35) N-Amerika")
25
Spartina towsendii 2n = 126 (n=63)
Voorbeeld van speciatie door hybridisatie op zeer korte termjn Spartina towsendii 2n = 126 (n=63)
")
26
ALLOPOLYPLOÏDIE 70 56 126 S. maritima S. x townsendii S alterniflora
AA AABB BB 70 56 126 ALLOPOLYPLOÏDIE

27
Polyploïde paren

28
Cardamine hirsuta Kleine veldkers 2n = 16 Akkers & braakland; nitrofiel Cardamine flexuosa Bosveldkers 2n = 32 Bossen & heggen; Vochtige bodem

29
Viola reichenbachiana
Donkersporig bosviooltje Viola riviniana Bleeksporig bosviooltje 2n = 20 2n = 40 A – VA Bossen, heggen , struwelen VA - VZ

30
Systematische praktijk:
Relatie tussen cytotype en afmeting van pollenkorrels

31
2n = 22 2n = 22 2n = 22 2n = 44 2n = 44 Conclusies uit de pollenmetingen: Andere variëteiten van G. capense allicht diploïd G. undulatum & G. chloroionanthum allicht polyploïd Het spurium complex is allicht een oude polyploïd (verlies van chromosomen) 2n = 44 2n = 44
 2n = 44. 2n = 44.")
32
Vorm & relatieve lengte:
Karyotype en idiogram

33
Karyosystematiek = microsystematiek ???

34
Variatie van het basisgetal in de Rosaceae s.l.
Rosoideae x = 7 (zelden 8 of 9) Prunoideae x = 8 Spiraeoideae x = 9 Pomoideae x = 17 Potentilla
 Prunoideae x = 8. Spiraeoideae x = 9. Pomoideae x = 17. Potentilla.")
35
Variatie van het basisgetal in de Rosaceae s.l.
Rosoideae x = 7 (zelden 8 of 9) Prunoideae x = 8 Spiraeoideae x = 9 Pomoideae x = 17 met monobasische reeksen van chromosoomgetallen met dibasische reeksen (17 = 8 + 9)
 Prunoideae x = 8. Spiraeoideae x = 9. Pomoideae x = 17. met monobasische reeksen van chromosoomgetallen. met dibasische reeksen. (17 = 8 + 9)")
36
Ranunculaceae Karyotypes op de achtergrond van de opvattingen in Natürlichen Pflanzenfamilien

37
Ranunculaceae Karyotypes op een beter onderbouwde fylogenie

38
1970 – “The New Karyosystematics”

Verwante presentaties



>")




 DoelgroepVerzondenOntvangen% LG wonen en dagbesteding.>")












