Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
formele analyse van taxonomische gegevens CLADISME (CLADISTIEK)
Van artificieel klasseren naar fylogenetische reconstructie Historische ontwikkeling van de systematiek 5. Ontwikkelingen in de twintigste eeuw formele analyse van taxonomische gegevens CLADISME (CLADISTIEK)
")
2
fylogenie = metafysica
Probleem evolutie = afgelopen historisch proces fylogenetisch onderzoek = historisch onderzoek testbaarheid hypothesen (sensu Popper) fylogenie = metafysica Fylogenie is metafysica, tenzij we over grote hoeveelheden fossielen zouden bezitten
 fylogenie = metafysica. Fylogenie is metafysica, tenzij we over grote hoeveelheden fossielen zouden bezitten.")
3
If organisms have reached their present state by a process of evolution,
it follows that they have a built‑in classification, and that man’s problem is to find it. This is quite a different problem from that presented to the classifier of man‑made objects ... R. Holttum (1967)
")
4
Willi Hennig 1950 Grundzüge einer Theorie der phylogenetischen Systematik

5
gemeenschappelijk bezit van afgeleide toestanden (synapomorfie)
basishypothese van de formele fylogenetische systematiek convergente (“parallelle”) kenmerken primitieve (plesiomorfe) toestanden afgeleide (apomorfe) toestanden gemeenschappelijk bezit van afgeleide toestanden (synapomorfie) = enige bewijs van afstammingsverwantschap
 kenmerken. primitieve (plesiomorfe) toestanden. afgeleide (apomorfe) toestanden. gemeenschappelijk bezit van afgeleide toestanden. (synapomorfie) = enige bewijs van afstammingsverwantschap.")
6
Een apomorfe toestand is niet noodzakelijk complexer dan een plesiomorge

7
Hennig, Grundzüge einer Theorie der phylogenetischen Systematik
grondbeginselen 1. ‘verwantschappen’ tussen levende en uitgestorven organismen zijn genealogisch 2. Fenotypische en genetische gelijkenissen zijn het resultaat van afstamming met modificatie (= evolutie) 3, cfr de basishypothese 3. Genealogische verwantschappen zijn te achterhalen via welbepaalde kenmerken die ze aantonen 4. Beste classificatie ≡ fylogenetische (genealogische) classificatie
 3, cfr de basishypothese. 3. Genealogische verwantschappen zijn te achterhalen via welbepaalde kenmerken die ze aantonen. 4. Beste classificatie ≡ fylogenetische (genealogische) classificatie.")
8
Cladistiek = studie van de transformatie van kenmerken voor het afleiden van en toetsen van fylogenetische hypothesen

10
Hennigiaanse argumentatie
Apomorfie 1 Plesiomorfie 0 Hennigiaanse argumentatie A B C a b c A B C d kenmerk d falsifieert hypothese afgeleid uit kenmerken a, b, c

11
A B C consensus cladogram

12
CLADOGRAM C is de buitengroep (‘outgroup) van A & B
taxa A en B zijn zustergroepen taxon A taxon B taxon C gemeenschappelijke voorouder (‘common ancestor’) van A en B CLADOGRAM
 van A en B. CLADOGRAM.")
13
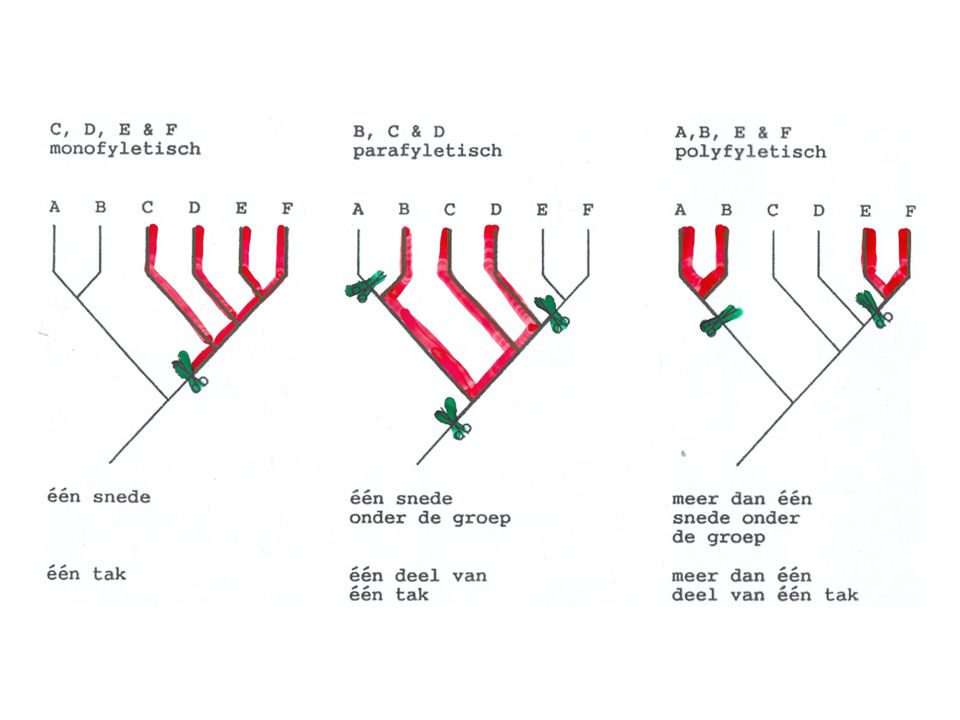
Cladisme ~ clade (monofyletische groep)
")
15
‘Reptilia’ zijn enkel monofyletisch door insluiting van de vogels

16
Werkwijze van de cladistiek
keuze studiegroep en EU’s (evolutionary units) criterium: beide monofyletisch!! 2. keuze kenmerken en toestanden 3. polariteitsbepaling 4. matrix met plesio- en apomorfieën 5. spaarzaamheidsanalyse Hennig 86, PAUP, PHYLIP 6. Vergelijking van de hypothesen (cladogrammen)
 criterium: beide monofyletisch!! 2. keuze kenmerken en toestanden. 3. polariteitsbepaling. 4. matrix met plesio- en apomorfieën. 5. spaarzaamheidsanalyse. Hennig 86, PAUP, PHYLIP. 6. Vergelijking van de hypothesen (cladogrammen)")
17
Werkwijze van de cladistiek

18
Polariteitsbepaling? buitengroepvergelijking Fossiele reeksen
Embryologische ontwikkeling ‘common equals primitive’

19
‘common equals primitive’
Apocarp = plesiomorfie van de angiospermen Apocarp = plesiomorfie van de Ranunculaceae Syncarp = plesiomorfie van de Solanaceae

20
Spaarzaamheidscriterium? (“parsimony”)
apomorfie plesiomorfie Spaarzaamheidscriterium? (“parsimony”) A B C A B C a b c d kenmerk d falsifieert hypothese afgeleid uit kenmerken a, b, c kenmerk d = “homoplasie”
 A B C. A B C. a. b. c. d. kenmerk d falsifieert hypothese afgeleid uit kenmerken a, b, c. kenmerk d = homoplasie")
22
Getransformeerde cladistiek
(Patterson, jaren 1980)
")
23
NUMERIEKE TAXONOMIE CLADISTIEK OTU: operational taxonomic unit
EU: evolutionary unit principieel fenetisch standpunt*: onderzoekt de fenetische verhoudingen tussen OTU’s * evolutie bestuderen zonder fossielen = metafysica (maar een goede fenetische classificatie weerspiegelt de evolutie) principieel fylogenetisch *: onderzoekt de cladistische verhoudingen tussen EU’s * de loop van de evolutie kan afgeleid worden uit kenmerken van hedendaagse organismen
 principieel fylogenetisch *: onderzoekt de cladistische verhoudingen tussen EU’s. * de loop van de evolutie kan afgeleid worden uit kenmerken van hedendaagse organismen.")
24
NUMERIEKE TAXONOMIE CLADISTIEK convergente kenmerken
OTU: operational taxonomic unit EU: evolutionary unit principieel fenetisch standpunt*: onderzoekt de fenetische verhoudingen tussen OTU’s principieel fylogenetisch *: onderzoekt de cladistische verhoudingen tussen EU’s convergente kenmerken plesiomorfieën & apomorfieën incl. autapomorfie (beïnvloedt fenetische afstand) convergente kenmerken enkel apomorfieën autapomorfieën zijn niet informatief multivariate analyse bij voorkeur > 100 kenmerken algemene statistische programma’s rekentijd kort bijzondere algoritmen enkele kenmerken volstaan reeds speciale programmatuur lange rekentijden fenogram (een dendrogram) in principe één hypothese cladogrammen meestal meer hypothesen synthetische classificatie, geen uitspraak over poly-, para- of monofyletische natuur van erkende groepen fylogenetische classificatie, enkel claden (monofyletische groepen) gevaar voor inflatie van aantal groepen
 convergente kenmerken. enkel apomorfieën. autapomorfieën zijn niet informatief. multivariate analyse. bij voorkeur > 100 kenmerken. algemene statistische programma’s. rekentijd kort. bijzondere algoritmen. enkele kenmerken volstaan reeds. speciale programmatuur. lange rekentijden. fenogram (een dendrogram) in principe één hypothese. cladogrammen. meestal meer hypothesen. synthetische classificatie, geen uitspraak over. poly-, para- of monofyletische natuur van erkende groepen. fylogenetische classificatie, enkel claden (monofyletische groepen) gevaar voor inflatie van aantal groepen.")
25
What does morphology tell us about orchid relationships?
(American Journal of Botany. 1999;86: ) © 1999 Botanical Society of America, Inc. What does morphology tell us about orchid relationships? — a cladistic analysis John V. Freudenstein and Finn N. Rasmussen A cladistic analysis of Orchidaceae was undertaken for 98 genera using 71 morphological apomorphies based on a reconsideration of previous character analyses and newly discovered variation. Een voorbeeld van een gepubliceerde cladistische analyse van morfologische gegevens
 © 1999 Botanical Society of America, Inc. What does morphology tell us about orchid relationships — a cladistic analysis. John V. Freudenstein and Finn N. Rasmussen. A cladistic analysis of Orchidaceae was undertaken for 98 genera using 71 morphological apomorphies based on a reconsideration of previous character analyses and newly discovered variation. Een voorbeeld van een gepubliceerde cladistische analyse van morfologische gegevens.")
26
All multistate characters were coded as unordered unless specified otherwise.
Character analysis 0. root tubers 0 = absent, 1 = present Root tubers are thickened roots that serve as organs of perennation, producing shoots in subsequent seasons. These structures occur primarily in the Diseae, Orchideae, and Diurideae and have been studied intensively (e.g., Irmisch, ; Germain de St.-Piérre, ; Prillieux, ; White, ; Ogura, ; Pridgeon and Chase, ). Triphora was scored as having a true root tuber, but Pogonia and Isotria were not, even though their unthickened roots are known to produce shoots (Ames, ). Nodular root thickenings are known from Apostasia (Stern and Warcup, ), Tropidia (Dressler, ), and possibly others, but these evidently do not produce new shoots. 1.root epidermis 0 = rhizodermis, 1 = velamen 2.exodermis 0 = unthickened, 1 = uniformly thickened, 2 = outer walls thickened 3.exodermal cell shape 0 = ± isodiametric, 1 = elongate
. Triphora was scored as having a true root tuber, but Pogonia and Isotria were not, even though their unthickened roots are known to produce shoots (Ames, 1922 ). Nodular root thickenings are known from Apostasia (Stern and Warcup, 1994 ), Tropidia (Dressler, 1981 ), and possibly others, but these evidently do not produce new shoots. 1.root epidermis 0 = rhizodermis, 1 = velamen. 2.exodermis 0 = unthickened, 1 = uniformly thickened, 2 = outer walls thickened. 3.exodermal cell shape 0 = ± isodiametric, 1 = elongate.")
27
48. seed laterally compressed walls 0 = absent, 1 =present
49. seed testa cell shape 0 = all isodiametric, 1 = end isodiametric, middle elongate, 2 = all elongate 50. seed striations 0 = absent, 1 = transverse/reticulate, 2 = longitudinal 51. seed intercellular spaces 0 = absent, 1 = present 52. seed wax caps 0 = absent, 1 = present 53. seed covered cell border 0 = absent, 1 = present Verklaar tegenspraak tekst (53 kenmerken) abstract (71 apomorfieën) 53 ↔ 71 ????
 abstract (71 apomorfieën) 53 ↔ 71")
28
The equally weighted analysis found most parsimonious trees with low consistency (CI = 0.29) but high retention (RI = 0.83). The strict consensus reveals a significant amount of structure, and most traditionally recognized subfamilies are supported as monophyletic, including the Apostasioideae, Cypripedioideae, Spiranthoideae, and Epidendroideae. Orchidoideae in the broad sense are paraphyletic, giving rise to spiranthoids. Vanilloids are sister to epidendroids, although exhibiting several states otherwise found only in clearly basal groups, such as Apostasioideae. The nonvandoid epidendroids are poorly resolved, due to a high degree of homoplasy. The vandoids appear to be monophyletic, contrary to recent molecular evidence, possibly due to repeated parallel development of the vandoid character suite.

29
Tick marks with numbers refer to clades discussed in the text.
Fig. 1. Consensus of most parsimonious trees from NONA analysis of equally weighted characters. Tick marks with numbers refer to clades discussed in the text. Decimal numbers are jackknife values (>0.50) for clades from replications. Jackknife analysis is similar to bootstrap analysis in that it is a way of testing the reliability of the dataset. It creates subsets of the original dataset by resampling contiguous sites of which their total number is smaller than that of the number of sites that make up the original dataset. This method allows you to analyse your dataset for a bias, which may be the result of the presence of, for instance, different domain stuctures in a single protein. Imagine that a protein has two similar domains that have been created by an event of gene duplication followed by the incorporation of a largely different number of mutations in the two domains due to adaptation to a new function or condition. One domain may have evolved into a regulatory domain, while the other domain kept its catalytic properties. Another example could be the case of multiple related genes such as the members of a gene family. Homologous recombination affecting only certain parts of such genes may lead in largely different apparent rates of evolution affecting different parts of such genes. Such differences in mutation rate may show up in a jackknife analysis when the subset size is set to 50% or less of the total size. Thus Jackknife analysis: is a statistical method for obtaining an estimate of error is used to evaluate the reliability of a tree
 for clades from replications. Jackknife analysis is similar to bootstrap analysis in that it is a way of testing the reliability of the dataset. It creates subsets of the original dataset by resampling contiguous sites of which their total number is smaller than that of the number of sites that make up the original dataset. This method allows you to analyse your dataset for a bias, which may be the result of the presence of, for instance, different domain stuctures in a single protein. Imagine that a protein has two similar domains that have been created by an event of gene duplication followed by the incorporation of a largely different number of mutations in the two domains due to adaptation to a new function or condition. One domain may have evolved into a regulatory domain, while the other domain kept its catalytic properties. Another example could be the case of multiple related genes such as the members of a gene family. Homologous recombination affecting only certain parts of such genes may lead in largely different apparent rates of evolution affecting different parts of such genes. Such differences in mutation rate may show up in a jackknife analysis when the subset size is set to 50% or less of the total size. Thus Jackknife analysis: is a statistical method for obtaining an estimate of error. is used to evaluate the reliability of a tree.")
30
Fig. 2. A single most parsimonious tree from the NONA analysis of equally weighted characters.
Black boxes are unique synapomorphies, and open boxes are homoplastic characters.

31
Figs. 4–7. Mapping of stamen characters on trees
from the present study and from Neyland and Urbatsch (1996a) 4. The most parsimonious optimization of the three stamen characters on the consensus tree from the present study. 5. An alternate, less parsimonious optimization on the consensus tree. 6–7. Two equally parsimonious optimizations of the three stamen characters on the tree of Neyland and Urbatsch (1996a : Fig. 2). Filled boxes are uniquely changing apomorphies, and open boxes are homoplastic characters. The number of the character is indicated above the box, and the number following the ">" under a box indicates the state to which that character changes.
 4. The most parsimonious optimization of the three stamen characters on the consensus tree from the present study. 5. An alternate, less parsimonious optimization on the consensus tree. 6–7. Two equally parsimonious optimizations of the three stamen characters on the tree of Neyland and Urbatsch (1996a : Fig. 2). Filled boxes are uniquely changing apomorphies, and open boxes are homoplastic characters. The number of the character is indicated above the box, and the number following the > under a box indicates the state to which that character changes.")
32
Voorbeeld = conventioneel concept

Verwante presentaties




![Deltion College Engels C1 Gesprekken voeren [Edu/002]/ subvaardigheid lezen thema: Order, order…. can-do : kan een bijeenkomst voorzitten © Anne Beeker.](/8/2048322/big_thumb.jpg "Deltion College Engels C1 Gesprekken voeren [Edu/002]/ subvaardigheid lezen thema: Order, order…. can-do : kan een bijeenkomst voorzitten © Anne Beeker.>")




to watch throughout the month of August,>")









