Download de presentatie
1
Eiwitten in het nieuws

2
Adenosine-receptor Nu 3D-structuur van drie receptoren van de 800 die we hebben… Belangrijk voor medicijnonderzoek (Parkinson, ….)
")
3
Green Fluorescent Protein
Nobelprijs Scheikunde 2008

5
Post-translationele modificaties Signals in sequences
Studietaak 3 Post-translationele modificaties Signals in sequences

6
Signals in sequences Eiwitten: meer dan alleen aminozuren
Post-translationele modificaties ‘Signaal’ hiervoor in az-volgorde! Voorspellingen over de structuur en/of functie van eiwitten Op basis van die signalen

7
Post-translationele modificaties

8
Post-translationele modificaties
Aanpassingen na afloop van (= ‘post’) translatie Extra ‘groepen’ die aan eiwitten worden gezet Invloed op o.m. vorm en functie van het eiwit Ken je zelf voorbeelden? Proteolytische klieving Protein sorting / protein targeting (pre-sequenties) Van inactief pro-eiwit naar actief eiwit (pro-sequenties)
 translatie. Extra ‘groepen’ die aan eiwitten worden gezet. Invloed op o.m. vorm en functie van het eiwit. Ken je zelf voorbeelden Proteolytische klieving. Protein sorting / protein targeting (pre-sequenties) Van inactief pro-eiwit naar actief eiwit (pro-sequenties)")
9
Signaaltransductie

10
EPO: geglycosileerd eiwit

11
Afsplitsing van aminozuren of peptiden

12
Afsplitsing van aminozuren of peptiden
Gebeurt door proteolytische enzymen Specifiek (herkenningssequentie)
")
13
Trypsine activeert andere zymogenen
Trypsine is de algemene activator van alle pancreatische zymogenen (pro-enzymen)
")
14
Clot composed of the protein fibrin

15
Elk organisme bezit proteolytische enzymen
Er zijn verschillende klasses proteolytische enzymen (ter info, niet kennen!) Allerlei functies Elk protease/proteïnase/peptidase heeft zijn eigen substraatspecificiteit = herkenningsplaats & klievingsplaats: alleen bepaalde aminozuurvolgordes! (straks meer)
 Allerlei functies. Elk protease/proteïnase/peptidase heeft zijn eigen substraatspecificiteit = herkenningsplaats & klievingsplaats: alleen bepaalde aminozuurvolgordes! (straks meer)")
16
Activatie van proteases
Een aantal proteases is betrokken bij het activeren van pro-eiwitten (zoals pro-insuline) Maar proteases worden zelf ook meestal gesynthetiseerd als pro-eiwitten Op de plaats van bestemming (bv. buiten de cel bij een bacterie of in het darmlumen bij de prancreatische proteases) wordt het pro-gedeelte afgesplitst (meestal door weer een ander protease) en wordt het protease actief
 Maar proteases worden zelf ook meestal gesynthetiseerd als pro-eiwitten. Op de plaats van bestemming (bv. buiten de cel bij een bacterie of in het darmlumen bij de prancreatische proteases) wordt het pro-gedeelte afgesplitst (meestal door weer een ander protease) en wordt het protease actief.")
17
Proteolytische enzymen in het biochemisch onderzoek
1: Bepaling van de aminozuursequentie van een eiwit

18
Bepaling van aminozuursequentie gebeurt tegenwoordig in 99,9% van de gevallen via DNA-technologie (HOE?), maar vroeger kon dat niet….
, maar vroeger kon dat niet….")
19
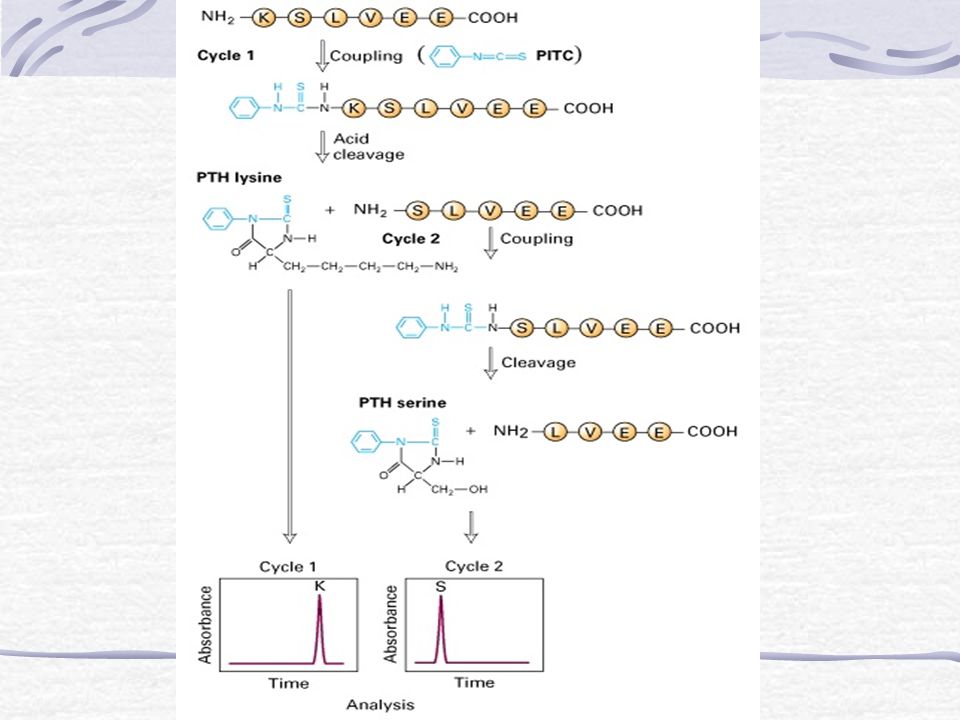
Ouderwetse bepaling van de az-sequentie
Pehr Edman ontwikkelde een methode voor het labelen van de N-terminus van een peptide, gevolgd door klieving ervan van het peptide Deze methode heet Edman-degradatie Door dit achter elkaar te doen en elke keer het gelabelde en vervolgens afgekliefde aminozuur te identificeren (mbv HPLC) kan de aminozuursequentie van peptiden tot ca. 50 az worden bepaald
 kan de aminozuursequentie van peptiden tot ca. 50 az worden bepaald.")
21
Ouderwetse bepaling van de az-sequentie
Een tweede manier is om vanaf de C-terminus steeds een aminozuur af te klieven m.b.v. carboxypeptidase en dit steeds te identificeren

22
Ouderwetse bepaling van de az-sequentie
Langere polypeptides moeten eerst in kleinere stukjes worden ´gehakt´ m.b.v. proteases of cyanogeenbromide (CNBr) Bijvoorbeeld: Enzym Klievingsplaats Trypsine Lys of Arg (C) Chymotrypsine Phe, Trp, Tyr (C) Pepsine Phe, Trp, Tyr (N) Cyanogenbromide Met (C)
 Bijvoorbeeld: Enzym. Klievingsplaats. Trypsine. Lys of Arg (C) Chymotrypsine. Phe, Trp, Tyr (C) Pepsine. Phe, Trp, Tyr (N) Cyanogenbromide. Met (C)")
23
Behandeling van een polypeptide met trypsine
Sub-peptides

24
N C (1) (2) Az-volgordes bepalen van de geproduceerde peptides
De az-volgorde van de sub-peptides is onbekend! De sequentie wordt gereconstrueerd d.m.b. de overlap tussen op verschillende manieren geproduceerde sub-peptides Trypsine (lys, arg) (1) Chymotrypsine (trp, tyr, phe) (2) N C Az-volgordes bepalen van de geproduceerde peptides
 (1) Chymotrypsine (trp, tyr, phe) (2) N. C. Az-volgordes bepalen van de geproduceerde peptides ")
25
Oefenen Ik heb een klein peptide van negen az
Na incubatie van dit peptide met trypsine (R K) vind ik twee peptides met de volgende az-volgordes: AAWGK TYVK Na incubatie van hetzelfde peptide met chymotrypsine (Y W F) vind ik een peptide met de volgende az-volgorde: VKAAW De overige brokstukken cq losse az van de incubatie met chymotrypsine zijn te klein om te isoleren en deze zijn dus niet bepaald! Wat is de az-volgorde van dit peptide?
 vind ik twee peptides met de volgende az-volgordes: AAWGK. TYVK. Na incubatie van hetzelfde peptide met chymotrypsine (Y W F) vind ik een peptide met de volgende az-volgorde: VKAAW. De overige brokstukken cq losse az van de incubatie met chymotrypsine zijn te klein om te isoleren en deze zijn dus niet bepaald! Wat is de az-volgorde van dit peptide")
26
Nog meer oefenen

27
Protein Targeting

28
Protein Targeting Bij prokaryoten zijn er niet zoveel verschillende locaties om een eiwit naar toe te sturen Welke? Eukaryote cellen hebben organellen en dus veel meer locaties om eiwitten naar toe te sturen

29
Prokaryoten: bekendste systeem: Sec-systeem

30
Protein Targeting bij eukaryoten
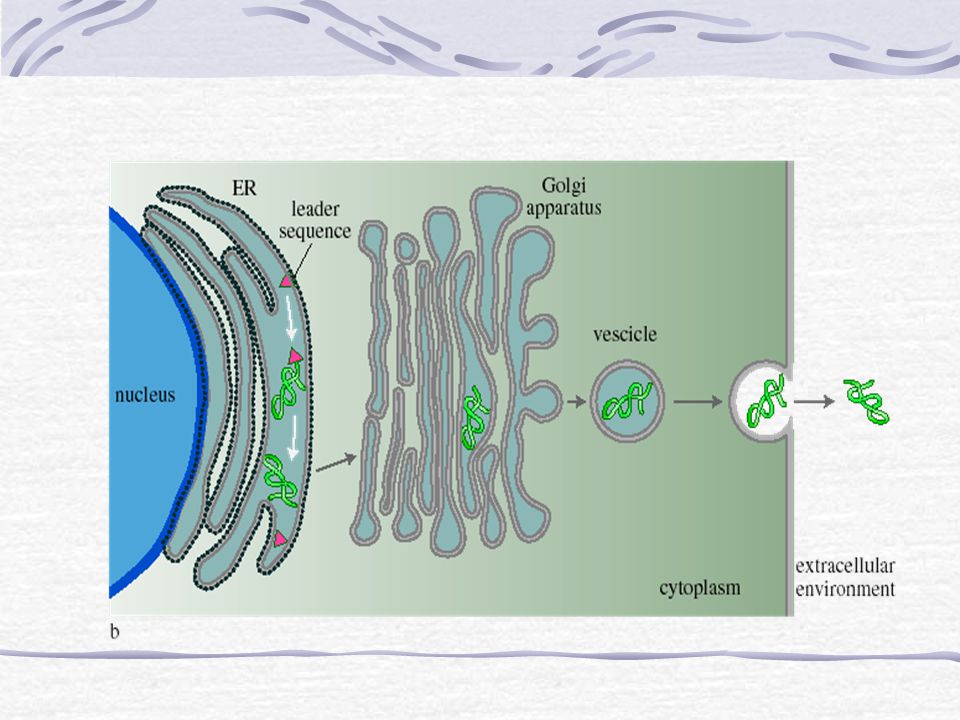
Twee ‘soorten’ ribosomen: Vrije ribosomen: maken eiwitten die in het cytosol zitten Aan ER gebonden ribosomen (‘ruw ER’) maken eiwitten voor endomembraansysteem ( organellen) en eiwitten die uitgescheiden worden Aan kernmembraan gebonden ribosomen maken eiwitten die in celkern terecht moeten komen Eiwitten voor endomembraansysteem of buitenkant: na synthese van N-terminaal signaalpeptide binding aan SRP (= signal-recognition particle) SRP met peptide en ribosoom aan SRP receptor op het ER; verwijdering van signaalpeptide
 maken eiwitten voor endomembraansysteem ( organellen) en eiwitten die uitgescheiden worden. Aan kernmembraan gebonden ribosomen maken eiwitten die in celkern terecht moeten komen. Eiwitten voor endomembraansysteem of buitenkant: na synthese van N-terminaal signaalpeptide binding aan SRP (= signal-recognition particle) SRP met peptide en ribosoom aan SRP receptor op het ER; verwijdering van signaalpeptide.")
31
Protein Targeting bij eukaryoten
Na koppeling ribosoom aan ER gaat synthese verder en eiwit komt in lumen ER terecht Daar blijft het of het wordt verder getransporteerd Zo zijn er ook signaalsequenties (‘postcodes’) voor mito’s, chloro’s, kern en andere organellen die geen deel uit maken van het endomembraan-systeem Hierbij wordt het eiwit eerst helemaal gemaakt in het cytosol en vervolgens naar het organel geïmporteerd
 voor mito’s, chloro’s, kern en andere organellen die geen deel uit maken van het endomembraan-systeem. Hierbij wordt het eiwit eerst helemaal gemaakt in het cytosol en vervolgens naar het organel geïmporteerd.")
34
om een goed overzicht over de verschillende modificaties te krijgen wordt nu eerst een proces behandeld, dat tijdens het hele modificatieproces voortduurt. 2 routes: Secretieeiwitten/membraan eiwitten en andere ER signaal sequentie (hydrophob) <->signaal recognation peptid (SRP) in ER opgevouwen -> de nieuw aangemaakt en goed opgevouwen eiwitten gaan via kleine secretie-korrels (vesicels) naar de Golgi waar ze evtl verder gemodificeerd worden Golgi bestaat uit een aantal schotelvormige ruimte (cisternae) cis en trans , verschillen van elkaar door de soorten enzymen. Sortering: via cis- en trans- Golgi naar hun plaats in de cel via vesicels Verschil tussen de routes, blauwe heeft geen ER signaal sequentie: posttranslationele translocatie
 <->signaal recognation peptid (SRP) in ER opgevouwen -> de nieuw aangemaakt en goed opgevouwen eiwitten gaan via kleine secretie-korrels (vesicels) naar de Golgi waar ze evtl verder gemodificeerd worden. Golgi bestaat uit een aantal schotelvormige ruimte (cisternae) cis en trans , verschillen van elkaar door de soorten enzymen. Sortering: via cis- en trans- Golgi naar hun plaats in de cel via vesicels. Verschil tussen de routes, blauwe heeft geen ER signaal sequentie: posttranslationele translocatie.")
37
Signaalsequenties De ‘postcodes’ van de cel Meer op

38
Extra groepen aan eiwitten

40
Extra groepen aan eiwitten
Glycosylering Acylering Myristoylering Methylering Fosforylering Sulfering Prenylering Hydroxylering Amidering Carboxylering …..

41
Glycosylering

42
. Een glycoproteïne is een eiwit met daaraan covalent gebonden een of meer koolhydraat-moleculen (ook wel glycanen genoemd). Het kan hierbij gaan om monosacharide(s), disacharide(s), oligosacharide(s), polysacharide(s) of afgeleiden hiervan (bv. sulfo- of fosfo-gesubstitueerd). Het kan gaan om één, enkele of veel koolhydraten per eiwit. Linkages besides 1-4 (eg 1-6, 1-2, 1-3) lead to branching of the sugar.
. Het kan hierbij gaan om monosacharide(s), disacharide(s), oligosacharide(s), polysacharide(s) of afgeleiden hiervan (bv. sulfo- of fosfo-gesubstitueerd). Het kan gaan om één, enkele of veel koolhydraten per eiwit. Linkages besides 1-4 (eg 1-6, 1-2, 1-3) lead to branching of the sugar.")
43
Hiervan is 90% N-linked glycanen
De meeste modificaties bij glycoproteïnen vinden plaats via N- of O-glycosidische bindingen O-linkage: Ser, Thr (GalNAc) N-linkage: Asn (GlcNAc) ~ 50% van alle eukaryote eiwitten is geglycosyleerd Hiervan is 90% N-linked glycanen Het vaakst worden de volgende monomere suikers aangetroffen: glucose, galactose, mannose, fucose, GalNAc en GlcNAc For proteins in the secretory pathway: a core glycan of Glc3Man9GlcNAc2 is added en bloc in the ER to proteins co-translationally (at the consensus N-X-S/T). As the protein traverses the Golgi this core glycan can be extensively modified, and O-linked glycans can be added. Description of glycosylation is made difficult by the COMPLEXITY of carbohydrate structures attached to the proteins: at least eight sugars, different glycosidic linkages.
 N-linkage: Asn (GlcNAc) ~ 50% van alle eukaryote eiwitten is geglycosyleerd. Hiervan is 90% N-linked glycanen. Het vaakst worden de volgende monomere suikers aangetroffen: glucose, galactose, mannose, fucose, GalNAc en GlcNAc. For proteins in the secretory pathway: a core glycan of Glc3Man9GlcNAc2 is added en bloc in the ER to proteins co-translationally (at the consensus N-X-S/T). As the protein traverses the Golgi this core glycan can be extensively modified, and O-linked glycans can be added. Description of glycosylation is made difficult by the COMPLEXITY of carbohydrate structures attached to the proteins: at least eight sugars, different glycosidic linkages.")
44
Eiwit-glycosylering vindt plaats tijdens het transport in ER en Golgi
Dus alleen eiwitten die via deze route gaan worden geglycosyleerd! (waar kun je dus geglycosyleerde eiwitten verwachten en waar niet??) De glycosylering is belangrijk voor de vouwing en de functie van het eiwit
 De glycosylering is belangrijk voor de vouwing en de functie van het eiwit.")
46
Cutis laxa: Een van de aandoeningen die het gevolg zijn van een glycosyleringsdefect

47
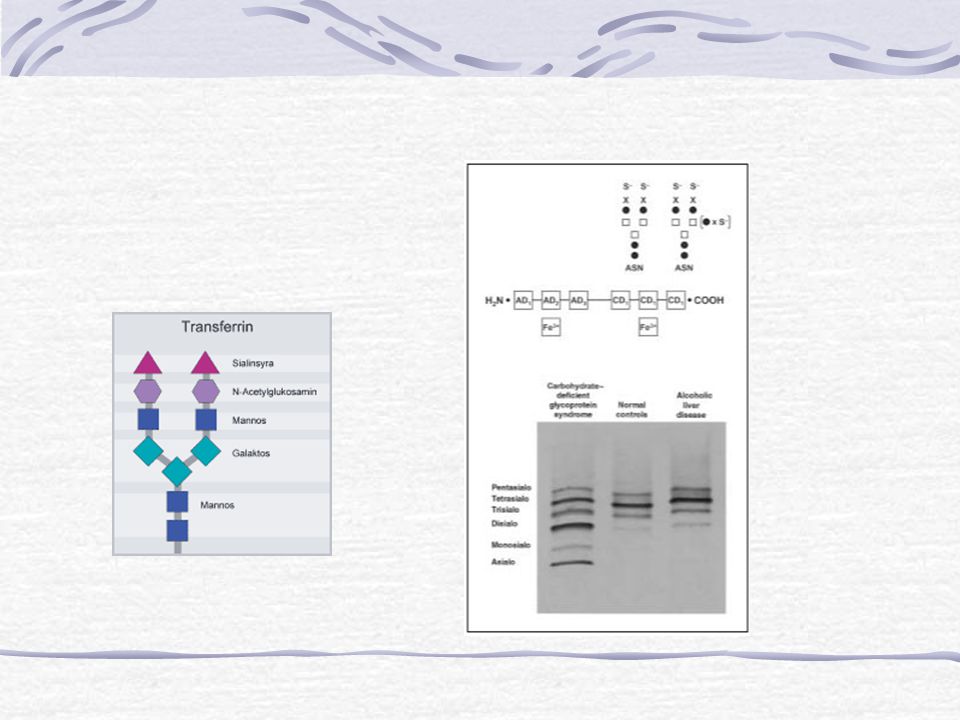
Hoe toon je aan of er bij een patiënt sprake is van een glycosyleringsdefect?
N-glycosylering: Isoëlektrische Focusering (IEF) van transferrine Transferrine in bloed Geglycosyleerd eiwit Afwezigheid of verstoring glycosylering a.g.v. algemeen glycsoyleringsdefect leidt tot afwijkend Isoëlektrisch Punt (IEP) van transferrine Sinds kort ook een klinische test voor O-glycosyleringsdefecten dankzij oud-studente Suzan Wopereis
 van transferrine. Transferrine in bloed. Geglycosyleerd eiwit. Afwezigheid of verstoring glycosylering a.g.v. algemeen glycsoyleringsdefect leidt tot afwijkend Isoëlektrisch Punt (IEP) van transferrine. Sinds kort ook een klinische test voor O-glycosyleringsdefecten dankzij oud-studente Suzan Wopereis.")
49
Voorspellingen Meer in de computerles

50
Voorspellingen Voorspellingen over de structuur en/of functie van eiwitten gebaseerd op: Vergelijken van eiwitten De aminozuurvolgorde van het eiwit zelf Signalen in aminozuursequenties

51
Voorspellende methodes
Van isoëlektrisch punt, via secundaire structuur en glycosyleringssites tot sorteersignalen Veel van deze methodes zijn te vinden bij het Expert Protein Analyis System Demo´s in computerles

52
Pas op! Voorspelling!!! Bijvoorbeeld N-glycosylering (N-X-(S,T)):
Programma ziet alle mogelijke N-glycosyleringssites Dit betekent NIET dat deze ook daadwerkelijk allemaal geglycosyleerd zijn Dat laatste kun je alleen in het lab aantonen






















.>")




