Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
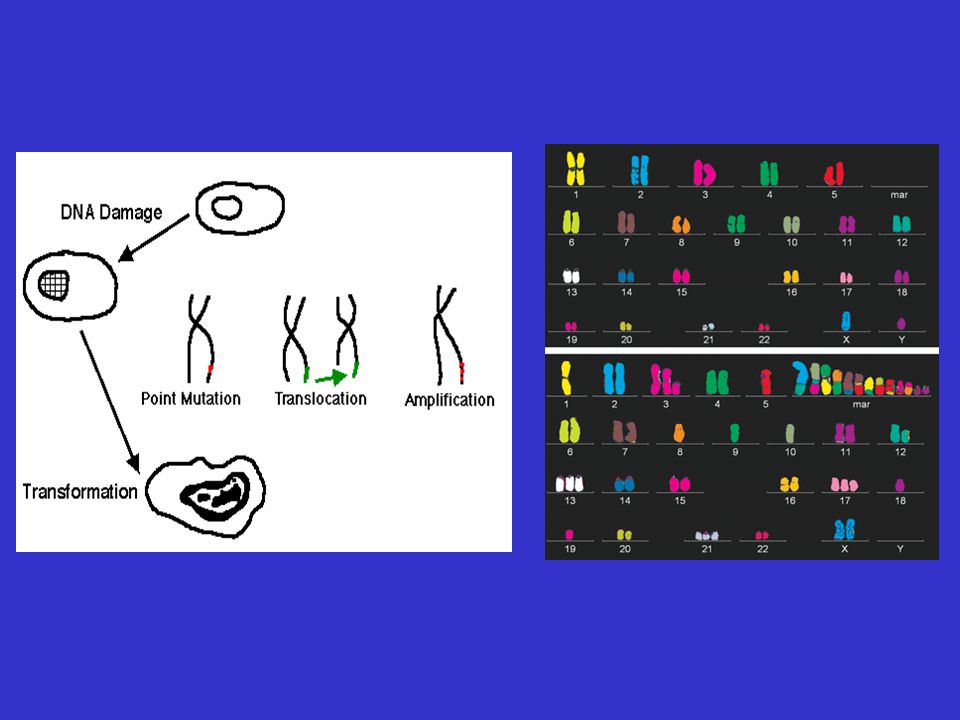
Ontstaan Neoplasieën ontstaan waarschijnlijk uit stamcellen of immature precursoren. De meeste zijn monoclonaal Men neemt aan dat een cel neoplastisch wordt door beschadiging van het DNA Voor de geboorte Beschadigd sperma of eicel Tijdens de embryonale ontwikkeling Na de geboorte Door een externe factor Tijdens het delen

2
= multifactoriëel Oorzaak van DNA beschadiging
“De”oorzaak van DNA beschadiging, die aanleiding geeft tot een neoplasie, bestaat niet; meestal bestaat deze DNA beschadiging uit meerdere afzonderlijke beschadigingen die elk het gevolg kunnen zijn van meerdere factoren = multifactoriëel Sommige van de factoren die in verband gebracht worden met het ontstaan van kanker kunnen we zelf controleren - (bvb omgevingsfactoren) - andere niet
 - andere niet.")
3
Figure 1: Age-adjusted incidence rates of the most common cancers
The Lancet, 360:861_868, 2002 Figure 1: Age-adjusted incidence rates of the most common cancers

4
HEPATOCELLULAIR CARCINOOM

5
Genen en kanker Poortwachter genen betrokken bij celgroei
Oncogenen Tumorsuppressorgenen Telomerase Opzichter genen betrokken bij DNA herstel o.a. Mismatch repair genen Bij afwijking: te veel of te weinig microsatelliet sequenties = Microsatelliet instabiliteit of MSI

7
Figure 1. Molecular pathways involved in tumour cell progression. (S.J. Diaz-Cano Histopathology 2008, 53, art.)
.")
8
Figure 2. Molecular pathways involved in microenvironment interactions (intercellular, and stromal, neoangiogenesis, and invasion) (S.J. Diaz-Cano Histopathology 2008, 53, art)
")
9
Figure 3. Molecular pathways involved in the tumour cell acquired capability of self-maintained proliferation. (S.J. Diaz-Cano Histopathology 2008, 53, art)
")
10
Figure 4. Molecular pathways involved in the tumour cell acquired capability of extended cell survival. (S.J. Diaz-Cano Histopathology 2008, 53, art)
")
11
Figure 5. Molecular pathways involved in the tumour cell acquired capability of genetic instability – DNA damage and repair (S.J. Diaz-Cano Histopathology 2008, 53, art)
")
12
Figure 6. Molecular pathways involved in the tumour cell acquired capability of modifications of chromatin, transcription and epigenetic changes. (S.J. Diaz-Cano Histopathology 2008, 53, art)
")
13
Figure 7. Molecular pathways involved in the tumour cell acquired capability of mobilization of cellular resources – ribosomes (protein synthesis). (S.J. Diaz-Cano Histopathology 2008, 53, )
")
14
Figure 8. Molecular pathways involved in the tumour cell acquired capability of mobilization of cellular resources – mitochondria (ATP synthesis, apoptosis, reactive oxygen species and damage). (S.J. Diaz-Cano Histopathology 2008, 53, art)
. (S.J. Diaz-Cano Histopathology 2008, 53, 1-19 art)")
15
Carcinogene factoren Chemisch Fysisch Biologisch

16
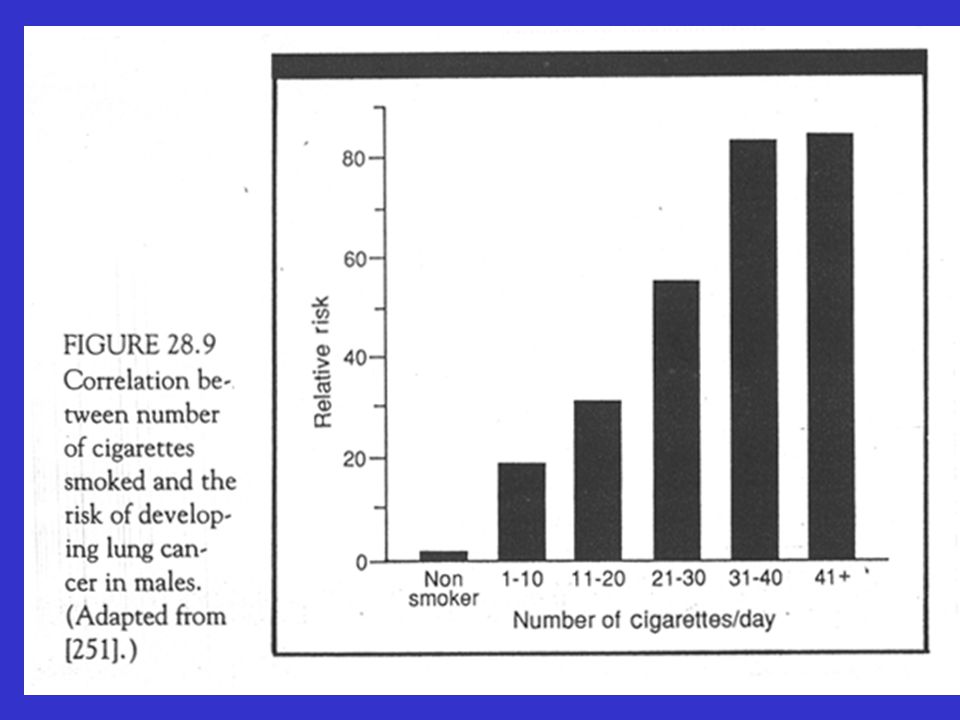
Chemische beroepsblootstelling Fysische factoren loniserende stralen :
Chemische factoren Genotsmiddelen Voeding Hormonen Chemische beroepsblootstelling Chemotherapie Fysische factoren UV stralen loniserende stralen : röntgen stralen radium medisch Hiroshima-Nagasaki Tsjernobyl Fysisch trauma Biologische factoren

17
Chemische beroepsblootstelling Fysische factoren loniserende stralen :
Chemische factoren Genotsmiddelen Voeding Hormonen Chemische beroepsblootstelling Chemotherapie Fysische factoren UV stralen loniserende stralen : röntgen stralen radium medisch Hiroshima-Nagasaki Tsjernobyl Fysisch trauma Biologische oorzaken van kanker

19
Chemische beroepsblootstelling Fysische factoren loniserende stralen :
Chemische factoren Genotsmiddelen Voeding Hormonen Chemische beroepsblootstelling Chemotherapie Fysische factoren UV stralen loniserende stralen : röntgen stralen radium medisch Hiroshima-Nagasaki Tsjernobyl Fysisch trauma Biologische oorzaken van kanker

22
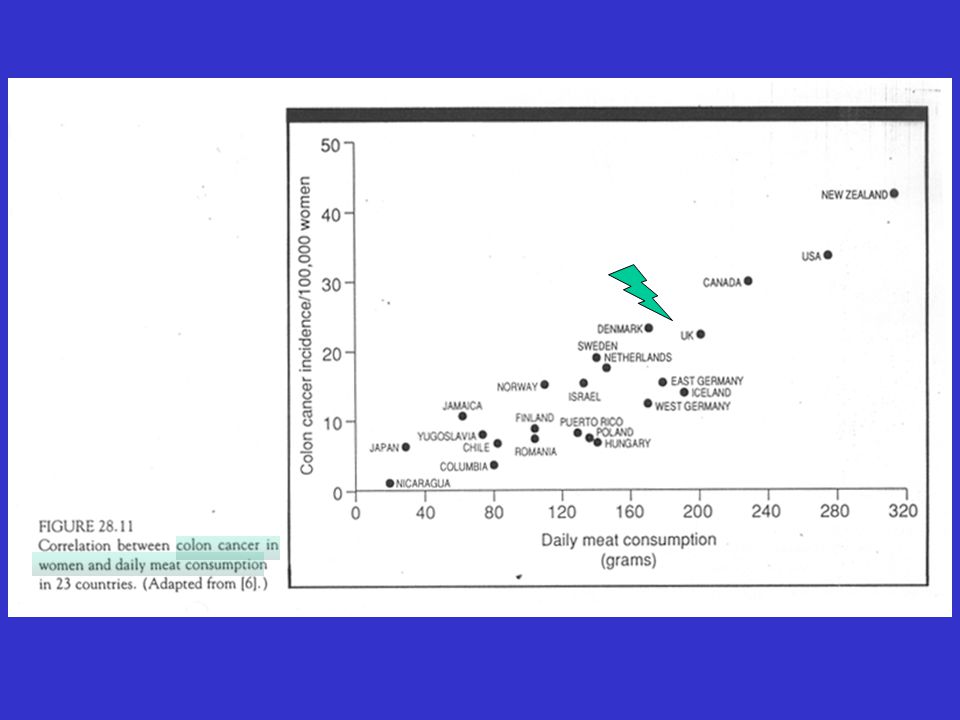
The Lancet, 360:861_868, 2002 Dietary risk factors, dietary protective factors, and other major risk factors for the common cancers Cancer Dietary and diet-related Dietary protective factors Other major risk factors risk factors Oral cavity, pharynx, Alcohol Probably fruit and vegetables Smoking and oesophagus Very hot drinks Obesity (adenocarcinoma of the oesophagus) Chinese-style salted fish (nasopharyngeal cancer) Stomach Probably high intake of salt- Probably fruit and vegetables Infection by Helicobacter pylori preserved foods and salt Colorectum Obesity Possibly red and processed Probably fruit, vegetables, and Sedentary lifestyle meat other plant foods rich in fibre Liver High alcohol intake None established Hepatitis viruses Foods contaminated with aflatoxins Pancreas None established None established Smoking Larynx Alcohol None established Smoking Lung None established Possibly fruit and vegetables Smoking Breast Obesity after menopause None established Reproductive and hormonal factors Alcohol Endometrium Obesity None established Low parity Cervix None established None established Human papillomavirus Prostate None established None established None established Kidney Obesity None established None established
 Chinese-style salted fish. (nasopharyngeal cancer) Stomach Probably high intake of salt- Probably fruit and vegetables Infection by Helicobacter pylori. preserved foods and salt. Colorectum Obesity. Possibly red and processed Probably fruit, vegetables, and Sedentary lifestyle. meat other plant foods rich in fibre. Liver High alcohol intake None established Hepatitis viruses. Foods contaminated with. aflatoxins. Pancreas None established None established Smoking. Larynx Alcohol None established Smoking. Lung None established Possibly fruit and vegetables Smoking. Breast Obesity after menopause None established Reproductive and hormonal factors. Alcohol. Endometrium Obesity None established Low parity. Cervix None established None established Human papillomavirus. Prostate None established None established None established. Kidney Obesity None established None established.")
23
Chemische beroepsblootstelling Fysische factoren loniserende stralen :
Chemische factoren Genotsmiddelen Voeding Hormonen Chemische beroepsblootstelling Chemotherapie Fysische factoren UV stralen loniserende stralen : röntgen stralen radium medisch Hiroshima-Nagasaki Tsjernobyl Fysisch trauma Biologische oorzaken van kanker

25
Tot hier

26
Chemische beroepsblootstelling Fysische factoren loniserende stralen :
Chemische factoren Genotsmiddelen Voeding Hormonen Chemische beroepsblootstelling Chemotherapie Fysische factoren UV stralen loniserende stralen : röntgen stralen radium medisch Hiroshima-Nagasaki Tsjernobyl Fysisch trauma Biologische oorzaken van kanker

27
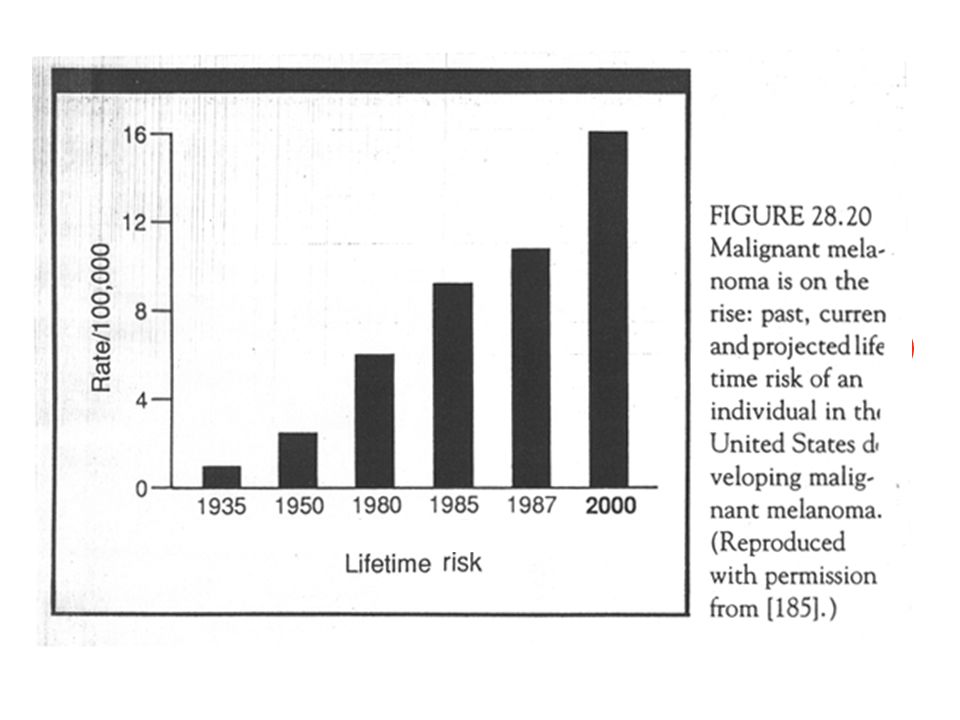
Maligne Melanoom

31
Chemische beroepsblootstelling Fysische factoren loniserende stralen :
Chemische factoren Genotsmiddelen Voeding Hormonen Chemische beroepsblootstelling Chemotherapie Fysische factoren UV stralen loniserende stralen : röntgen stralen radium medisch Hiroshima-Nagasaki Tsjernobyl Fysisch trauma Biologische oorzaken van kanker

32
VIRUSSEN Virchows Archiv maart 2000

33
Biologische oorzaken van kanker
Bacteriën HP: Maaglymfoom Maagadenocarcinoom Parasieten Schistosoma Blaas: plaveiselcelcarcinoom

34
Papilloom van het plaveiselepitheel (HPV)
")
35
HPV - geassocieerde papillomen

36
Verruca vulgaris

37
TUMOREN - Definities - Ontstaan & evolutie Oorzaken Precancereuze condities Metastasering - Verdediging - Labo-technieken (Morfologische diagnose)
")
38
carcinogenese KANKERCEL NORMALE CEL ONCOGEN TUMOR SUPPRESSORGEN
ACTIVATIE DESACTIVATIE RECESSIEF DOMINANT NORMALE CEL KANKERCEL Ongecontroleerde groei Gecontroleerde groei

39
-turn on oncogenen (groeipromotie) -turn off tumorsupressor genen( verlies groei-inhibitie) neoplasie
 -turn off tumorsupressor genen( verlies groei-inhibitie) neoplasie")
40
Oncogenen Groeifactoren Groeifactor receptoren
c-sis gen Groeifactor receptoren C-erb-b2 (neu) Cyclische nucleotide bindende eiwitten HRAS KRAS NRAS Tyrosine fosforylerende kinases ABL SRC Nucleaire transcryptiefactoren MYC
 Cyclische nucleotide bindende eiwitten. HRAS. KRAS. NRAS. Tyrosine fosforylerende kinases. ABL. SRC. Nucleaire transcryptiefactoren. MYC.")
41
Tumor suppressorgenen
RB P53 NF VHL APC ………

42
Telomerase Telomeren Repetitieve sequenties van niet coderend DNA aan uiteinde van de chromosomen Bij elke celdeling gaat stukje telomeer DNA verloren Eens het telomeer DNA te kort is kan geen celdeling meer plaatsgrijpen Te korte telomeren worden hersteld door telomerase, maar telomerase komt fysiologisch enkel voor in stamcellen en germinatieve cellen. In grote meerderheid van tumoren is telomerase geactiveerd

43
Late chromosomale veranderingen
Door ontregeling van de celdeling kunnen uiteindelijk cellen ontstaan waarbij volledige chromosomen of grote stukken chromosomen ontbreken of vermenigvuldigen

44
Virchows arch 2004, 444: Negative and positive regulators of the normal cell cycle. Signals promoting and inhibiting the different phases of the cell cycle as well as checkpoints monitoring the proper completion of every phase of the cell cycle are indicated. In the centre of the cycle, the CDK/cyclin complexes driving the respective phase are shown. For details, see the text

45
Virchows arch 2004, 444:131-323 Upregulation of cell-cycle activators
downregulation of cell-cycle inhibitors Deregulation of the cell cycle in cancer. Upregulation of cell-cycle activators and downregulation of cell-cycle inhibitors are both involved in promoting the transformation of a normal cell into a continuously proliferating cell, which is independent of growth-promoting signals and resistant to growth-inhibiting signals. When this transformation is supported by other mechanisms, such as angiogenesis as well as evasion of apoptosis and immune surveillance, it will create the clonogenic malignant cell. For every alteration of cell-cycle regulators, only one example of an associated human malignancy is given. The two-dimensional representation is clearly simplifying the complex interdependence of all participating factors. For further details, see the text and references therein

46
Nature, vol 432, 18 nov 2004: Figure 4 Schematic representation of two main steps that contribute to a spectrum of mutations leading to cancer development. If DNA damage is repaired efficiently, the likelihood of tumour development is low. If cells have mutations in DNA-damage-response signalling pathways — either sporadic or inherited — this will lead to enhanced genomic abnormalities. Cells with damaged DNA frequently arrest or do not survive, thus reducing the probability that they will progress to malignancy. Mutations in apoptosis pathways, DNA-damage, DNA-repair or mitotic-checkpoint pathways can permit the survival or continued growth of cells with genomic abnormalities, thus enhancing the likelihood of malignant transformation.

47
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
48
TUMOREN - Definities - Ontstaan & evolutie Oorzaken Precancereuze condities Metastasering - Verdediging - Labo-technieken (Morfologische diagnose)
")
49
Precancereuze condities
1) Genetische predispositie : in sommige families komt kanker méér voor dan in andere. 2) Afwijkingen in het orgaan zelf : thv sommige letsels ontwikkelt zich meer frequent een neoplasie; nl thv - metaplasie - dysplasie - GA tumoren - Chronische inflammatie
 Genetische predispositie : in sommige families komt kanker méér voor dan in andere. 2) Afwijkingen in het orgaan zelf : thv sommige. letsels ontwikkelt zich meer frequent een. neoplasie; nl thv. - metaplasie. - dysplasie. - GA tumoren. - Chronische inflammatie.")
50
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
51
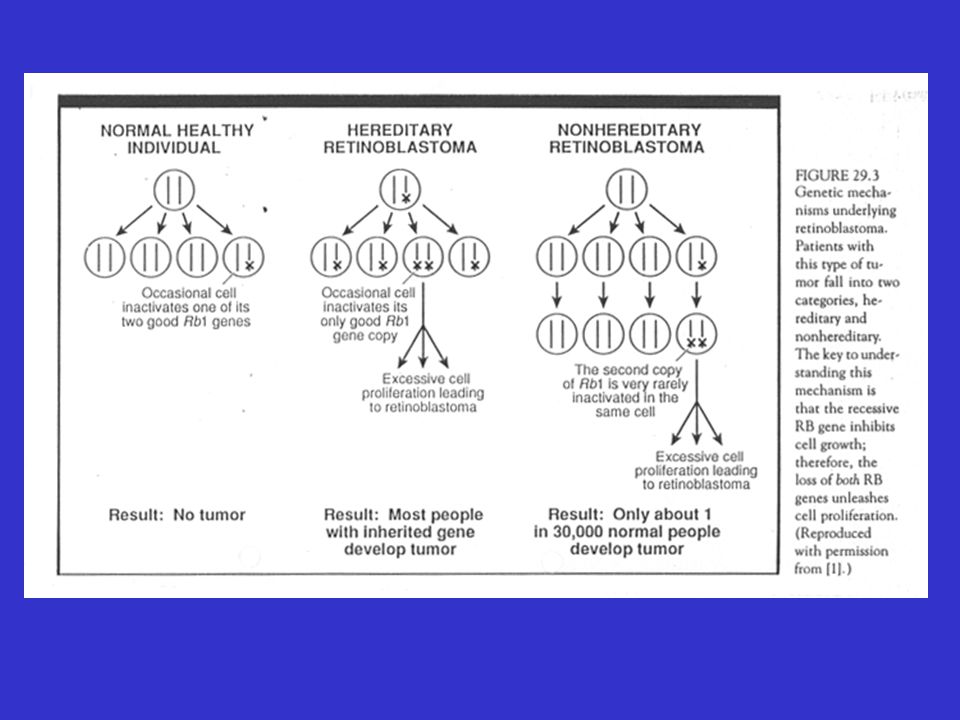
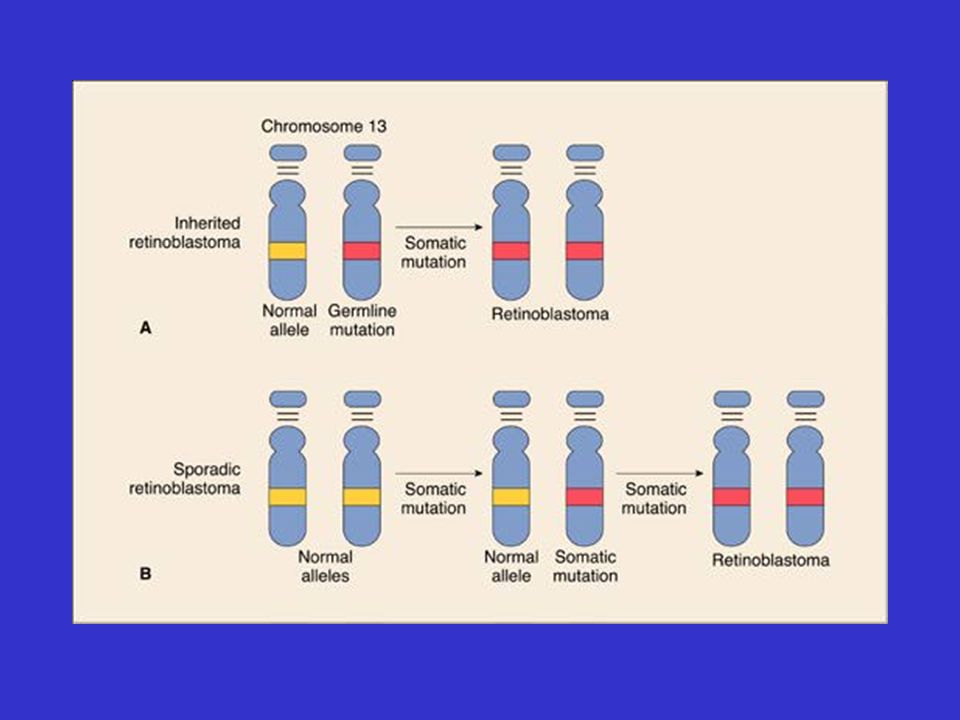
Genetische predispositie
Gevolg van mutatie thv een tumorsuppressorgen Het bestaan van tumor-suppressorgenen werd vermoed door het bestuderen van de tumor: “retinoblastoom” 2 vormen : hereditair en sporadisch hereditaire vorm: jonge leeftijd; kan bilateraal sporadische vorm: volwassen leeftijd;unilateraal

52
Retinoblastoom

53
RETINOBLASTOOM

56
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
58
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
59
Poliepen van de darm

60
Adenomateuze poliposis coli - M-13jaar

61
Syndrome Tumour caused Defect
Germ-line mutaties in tumorsuppressorgenen Syndrome Tumour caused Defect MEN syndromes Multiple tumours in endocrine organs Mutations on chromosomes 10 and 11 Polyposis coli (APC) Adenomata and carcinomas of the colon Absent tumour suppressor gene Li-Fraumeni (P53) Breast cancer and sarcomas Mutated tumour suppressor gene Familial retinoblastoma (RB) Malignant tumour of the retina Neurofibromatosis type 1 (NF) Benign and malignant tumours of peripheral nerves Abnormal tumour suppressor gene
 Adenomata and carcinomas of the colon. Absent tumour suppressor gene. Li-Fraumeni. (P53) Breast cancer and sarcomas. Mutated tumour suppressor gene. Familial retinoblastoma (RB) Malignant tumour of the retina. Neurofibromatosis type 1 (NF) Benign and malignant tumours of peripheral nerves. Abnormal tumour suppressor gene.")
62
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
63
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocation of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocation of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
64
C-erb-B2 oncogen werkt via vermenigvuldigen van het normaal proto-oncogen; dit leidt tot de overproductie van een proteine dat ongecontroleerde celgroei veroorzaakt. Dit proteine wordt gedetecteerd door een immunoperoxidase kleuring; c-erb-B2 immuunhistochemie in borst carcinoma

65
Oncogenesis= afwijking genoom
Mechanism Action Example Loss of Tumor Suppressor Gene Function Loss of normal growth inhibition BRCA-1 Lack of regulation of cell adhesion with loss of growth control through cell interaction APC Loss of down-regulation of growth promoting signal transduction NF-1 Loss of regulation of cell cycle activation through sequestration of transcriptional factors Rb Loss of regulation of cell cycle activation through lack of inhibition of cell proliferation that allows DNA repair p53 Growth Promotion Overexpression of growth factor receptors (such as epidermal growth factor, or EGF) making cells more sensitive to growth stimuli c-erb-B2 Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth ras Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited c-abl Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle c-myc Limitation of Apoptosis Overexpression of gene, activated by translocation prevents apoptosis bcl-2
 making cells more sensitive to growth stimuli. c-erb-B2. Increased growth factor signal transduction by an oncogene that lacks the GTPase activity that limits GTP induction of cytoplasmic kinases that drive cell growth. ras. Overexpression of a gene product by stimulation from an oncogene (such as ras) c-sis. Lack of normal gene regulation through translocatiion of a gene where it is controlled by surrounding gene to a place where it is no longer inhibited. c-abl. Binding of oncogene product to the nucleus with DNA transcriptional activation to promote entry into the cell cycle. c-myc. Limitation of Apoptosis. Overexpression of gene, activated by translocation prevents apoptosis. bcl-2.")
66
bcl-2 oncogen overexpressie leidt tot inhibitie van apoptose en vermenigvuldiging van de lymfocyten. De tumorale lymfocyten in de lymphoiede follikels en de interfollicular gebieden worden gekleurd. Immuunhistochemie voor bcl-2

67
Precancereuze condities
1) Genetische predispositie : in sommige families komt kanker méér voor dan in andere. 2) Afwijkingen in het orgaan zelf : thv sommige letsels ontwikkelt zich meer frequent een neoplasie; nl thv - metaplasie - dysplasie - GA tumoren - Chronische inflammatie
 Genetische predispositie : in sommige families komt kanker méér voor dan in andere. 2) Afwijkingen in het orgaan zelf : thv sommige. letsels ontwikkelt zich meer frequent een. neoplasie; nl thv. - metaplasie. - dysplasie. - GA tumoren. - Chronische inflammatie.")
68
Precancereuze letsels
1. Metaplasie 2. Dysplasie 3. Goedaardige tumoren 4. Chronische inflammatie

69
Metaplasie Definitie :
Vervanging van de cel door een cel met een andere (normaal) fenotype die in een andere localisatie thuishoort
 fenotype die in een andere localisatie thuishoort.")
70
Metaplasie Oorzaken : chronische inflammatie = meest frequent
- Respiratoir : pseudo-meerlagig trilhaarepitheel vervangen door plaveiselepitheel bij chronische bronchitis - maagepitheel vervangen door dundarm epitheel bij chronische gastritis.

71
Nomenclatuur metaplasie
1) Het vervangend weefsel wordt genoemd gevolgd door het woord “metaplasie” gevolgd door het orgaan, vb. : plaveisel-metaplasie van de bronchus, intestinale-metaplasie van de maag, kraakbeen-metaplasie van de long. 2) Meer specifiek : leukoplakie = macroscopisch begrip = wit worden van weefsel dat normaal niet wit is. - plaveisel-metaplasie van een cylinderepitheel (vb. cervix mucosa) - verhoorning-metaplasie van niet-verhoornd plaveiselepitheel (vb. mond mucosa) Barrett oesofagus : maag- of intestinale metaplasie van de distale oesofagus wordt Barrett oesofagus genoemd.
 Het vervangend weefsel wordt genoemd gevolgd door het woord metaplasie gevolgd door het orgaan, vb. : plaveisel-metaplasie van de bronchus, intestinale-metaplasie van de maag, kraakbeen-metaplasie van de long. 2) Meer specifiek : leukoplakie = macroscopisch begrip = wit worden van weefsel dat normaal niet wit is. - plaveisel-metaplasie van een cylinderepitheel (vb. cervix mucosa) - verhoorning-metaplasie van niet-verhoornd plaveiselepitheel (vb. mond mucosa) Barrett oesofagus : maag- of intestinale metaplasie van de distale oesofagus wordt Barrett oesofagus genoemd.")
72
Kliniek metaplasie Meestal weinig gevolgen – kan dit letsel CA geven ? Het is het gevolg van celbeschadiging en vele kankers zijn dit eveneens. De ontwikkeling van kanker kan dus door een stadium van metaplasie gaan

73
Precancereuze letsels
1. Metaplasie 2. Dysplasie 3. Goedaardige tumoren 4. Chronische inflammatie

74
Dysplasie Definitie : Abnormale schikking en oriëntatie van cellen, gepaard met cel- en kernafwijkingen en een toegenomen aantal mitosen.

75
Dysplasie: Meestal tgv chronische irritatie en inflammatie
gepaard met metaplasie volgend op metaplasie de novo

76
Morfologie dysplasie :
1. Kern/plasma (K/P) verhouding stijgt. 2. Hyperchromatose van de kern en/of grote nucleolen en/of verdikte chromatine. 3. Neiging tot anaplasie. Anaplasie = verminderen van de differentiatie, bvb. verlies aan kenmerken eigen aan de cel (vb. mucusproductie, afplatting van bovenste cellen in plaveiselepitheel, verlies paraplucellen). 4. Mitosen op abno plaats bvb: hoger dan basale cellaag in plaveiselepitheel hoger dan onderste 1/2 crypten in colon
 verhouding stijgt. 2. Hyperchromatose van de kern en/of grote. nucleolen en/of verdikte chromatine. 3. Neiging tot anaplasie. Anaplasie = verminderen van de differentiatie, bvb. verlies aan kenmerken eigen aan de cel. (vb. mucusproductie, afplatting van bovenste. cellen in plaveiselepitheel, verlies paraplucellen). 4. Mitosen op abno plaats. bvb: hoger dan basale cellaag in plaveiselepitheel. hoger dan onderste 1/2 crypten in colon.")
77
Gradering : * Lichte dysplasie * Matige dysplasie
* Zware dysplasie of carcinoma in situ (C.I.S) = maligne letsel C.I.S. wordt gebruikt als synoniem voor zware dysplasie van een epitheel. Men spreekt ook van intra-epitheliale neoplasie, Nu ook gebruikt voor lichte en matige dysplasie: bvb. : CIN = cervical intra-epithelial neoplasia. CIN I = lichte dysplasie CIN II = matige dysplasie CIN III = zware dysplasie of CIS Naar analogie wordt deze naamgeving ook voor andere organen gebruikt (prostaat=PIN, vagina=VIN enz...) Laatste evolutie: laagradig (I) en hooggradig (II & III)
 = maligne letsel. C.I.S. wordt gebruikt als synoniem voor zware dysplasie van een epitheel. Men spreekt ook van intra-epitheliale neoplasie, Nu ook gebruikt voor lichte en matige dysplasie: bvb. : CIN = cervical intra-epithelial neoplasia. CIN I = lichte dysplasie. CIN II = matige dysplasie. CIN III = zware dysplasie of CIS. Naar analogie wordt deze naamgeving ook voor andere organen gebruikt (prostaat=PIN, vagina=VIN enz...) Laatste evolutie: laagradig (I) en hooggradig (II & III)")
78
Dysplasie cervix epitheel

79
Dysplasie cervix epitheel

80
Dysplasie cervix epitheel

81
Plaveiselepitheel Normaal Dysplasie

82
Plaveiselepitheel dysplasie

83
Plaveiselepitheel dysplasie

84
Carcinoma in situ

85
Invasief carcinoom cervix

86
Invasief carcinoom cervix

87
Invasief carcinoom cervix

88
Invasief carcinoom cervix

89
Invasief carcinoom cervix

90
Overgang naar kanker bij darmepitheel

91
Normaal epitheel / dysplastisch epitheel

92
Klierepitheel met lichte dysplasie

93
Adenoom darm matige dysplasie epitheel

94
Klierepitheel met matige dysplasie

95
Klierepitheel:zware dysplasie

96
Klierepitheel: zware dysplasie

97
Klierepitheel zware dysplasie

98
Invasief adenocarcinoom colon

99
Precancereuze letsels
1. Metaplasie 2. Dysplasie 3. Goedaardige tumoren 4. Chronische inflammatie

100
Chronische inflammatie
Vooral toen er geen behandeling was voor chronische ontsteking Lupus vulgaris Osteomyelitis Chronische gastritis Levercirrose RCUH

101
Nature, vol 432, 18 nov 2004: Figure 2 Model for carcinogenesis resulting from persistence of a state of injury repair. A, Cellular events of epithelial repair. a, Resting epithelium with several differentiated cell phenotypes (brown, orange, and yellow) derived from tissue stem cells, now quiescent (red). Pathways such as Hh and Wnt signalling pathways that have a role in the renewal of stem cells are not active. b, Epithelial defect resulting from acute injury. Loss of epithelial continuity activates a repair program which is driven by Hh or Wnt signalling. This program results in the acquisition by epithelial cells of a more mesenchymal phenotype, including flattening and movement of cells (straight arrow) to cover the wound, activation (green), and expansion of stem cells through renewal divisions (curved arrows). c, The wound is repaired, first by rapid cell movement, and then by restoration of cell numbers resulting from the amplification of stem cells and the differentiation of their progeny. Subsequently, either epithelial continuity and patterning is restored, Hh and Wnt signalling ceases, and the stem cell compartment returns to quiescence (a); or oncogenic event(s) may trap a stem cell in an activated state of continuous renewal, which is driven by autonomous Wnt or Hh signalling (d). Further genetic or epigenetic change in such a persistently activated stem cell (curved red arrows) might produce a cancer stem cell (green) which is capable of aggressively propagating a cancer (e). This may result from enhanced proliferation and production of more cancer stem cells as well as from differentiated cancer cells (blue). B, Stem cells cycle between quiescence and activity as a consequence of Hh/Wnt driven responses to injury. Oncogenic event(s) may trap activated stem cells in a permanent state of Hh/Wnt driven activity, resulting in cancer stem cells.
 derived from tissue stem cells, now quiescent (red). Pathways such as Hh and Wnt signalling pathways that have a role in the renewal of stem cells are not active. b, Epithelial defect resulting from acute injury. Loss of epithelial continuity activates a repair program which is driven by Hh or Wnt signalling. This program results in the acquisition by epithelial cells of a more mesenchymal phenotype, including flattening and movement of cells (straight arrow) to cover the wound, activation (green), and expansion of stem cells through renewal divisions (curved arrows). c, The wound is repaired, first by rapid cell movement, and then by restoration of cell numbers resulting from the amplification of stem cells and the differentiation of their progeny. Subsequently, either epithelial continuity and patterning is restored, Hh and Wnt signalling ceases, and the stem cell compartment returns to quiescence (a); or oncogenic event(s) may trap a stem cell in an activated state of continuous renewal, which is driven by autonomous Wnt or Hh signalling (d). Further genetic or epigenetic change in such a persistently activated stem cell (curved red arrows) might produce a cancer stem cell (green) which is capable of aggressively propagating a cancer (e). This may result from enhanced proliferation and production of more cancer stem cells as well as from differentiated cancer cells (blue). B, Stem cells cycle between quiescence and activity as a consequence of Hh/Wnt driven responses to injury. Oncogenic event(s) may trap activated stem cells in a permanent state of Hh/Wnt driven activity, resulting in cancer stem cells.")
102
TUMOREN - Definities - Ontstaan & evolutie Oorzaken
Precancereuze condities Metastasering - Verdediging - Morfologische diagnose (labotechnieken)
")
103
Metastasiëring: theorieën Clonale selectie
Gepredetermineerde tumorcellen Gastheer factoren Lancet 2007; 369

104
Lancet 2007; 369

105
Lancet 2007; 369

106
Lancet 2007; 369 The process of metastasis begins before cells migrate from a primary tumour mass. Among the earliest characteristics of transformed cells are genetic and phenotypic instability. Cancer cells are more prone to mutation and phenotypic drift than their normal counterparts. Genetic instability, coupled with a Darwinian type of selection—survival of the fittest—results in populations resistant to normal homoeostatic growth controls, immune attack, and environmental restraints.6 The rate of progression varies and, within any neoplastic mass, subpopulations can be isolated with different malignant potential. Thus, not all tumours are metastatic, nor are all cells within so-called metastatic tumours capable of metastasising. Even cells isolated from large metastases show substantial heterogeneity when assessed experimentally, raising questions as to whether cells might transiently acquire metastatic potential. Recent evidence suggests that tumour cells might begin conditioning distant tissues for colonisation by establishing a so-called pre-metastatic niche. As yet unknown factors mobilise haematopoietic stem cells to tissues, remodel the matrix, and modify stromal cells and the growth factor milieu such that tumour cells are attracted to or have increased predilection for growth at these sites. In a transgenic mouse colon carcinoma model, CD34+ immature myeloid cells expressing the chemokine receptor CCR1 were recruited from the bone marrow to the edges of local primary lesions and stimulated local invasion by tumour cells expressing the ligand CCL9. Importantly, genetic instability, generation of variants, and establishment of pre-metastatic niches represent intrinsic tumour cell and microenvironmental changes that take place before cancer cell dissemination.

107
Lancet 2007; 369 Epithelial-mesenchymal transition
Neoplastic cells might acquire the ability to metastasise by dedifferentiation to a more motile mesenchymal cell phenotype, a process called epithelial-mesenchymal transition (EMT). Once established in a new environment, metastatic cells might then revert back to a non-metastatic phenotype, via a mesenchymal-epithelial transition. Epithelial-mesenchymal transition can be induced by different stimuli, with transforming growth factor (TGF) signalling having a key role. Other important mediators include oncogenic signalling pathways (notably phosphoinositide 3 [PI3] kinase), mitogen-activated protein (MAP) kinases, loss of E-cadherin (or a switch to N-cadherin), and activation of transcription regulators such as Twist and Snail (SNA1). Interestingly, Wnt, Notch, and Hedgehog signalling pathways (also implicated in stem cell maintenance) are linked to epithelial-mesenchymal transition. Cells induced to undergo epithelial-mesenchymal transition not only exhibit enhanced motility but also are resistant to apoptosis: key requirements for successful metastasis. However, other cancer cells might use a collective migration that is independent of an epithelial-mesenchymal transition. The fact that Wnt signalling can also induce collective migration in addition to epithelial-mesenchymal transition emphasises the complex interrelations and plasticity in all of these processes. Although a role for epithelial-mesenchymal transition during development is well accepted and can be demonstrated and manipulated in many experimental tumour models, some question whether it occurs in human cancers, and it is important to state explicitly that epithelial-mesenchymal transition is not synonymous with invasion or metastasis.
. Once established in a new environment, metastatic cells might then revert back to a non-metastatic phenotype, via a mesenchymal-epithelial transition. Epithelial-mesenchymal transition can be induced by different stimuli, with transforming growth factor (TGF) signalling having a key role. Other important mediators include oncogenic signalling pathways (notably phosphoinositide 3 [PI3] kinase), mitogen-activated protein (MAP) kinases, loss of E-cadherin (or a switch to N-cadherin), and activation of transcription regulators such as Twist and Snail (SNA1). Interestingly, Wnt, Notch, and Hedgehog signalling pathways (also implicated in stem cell maintenance) are linked to epithelial-mesenchymal transition. Cells induced to undergo epithelial-mesenchymal transition not only exhibit enhanced motility but also are resistant to apoptosis: key requirements for successful metastasis. However, other cancer cells might use a collective migration that is independent of an epithelial-mesenchymal transition. The fact that Wnt signalling can also induce collective migration in addition to epithelial-mesenchymal transition emphasises the complex interrelations and plasticity in all of these processes. Although a role for epithelial-mesenchymal transition during development is well accepted and can be demonstrated and manipulated in many experimental tumour models, some question whether it occurs in human cancers, and it is important to state explicitly that epithelial-mesenchymal transition is not synonymous with invasion or metastasis.")
108
Lancet 2007; 369 Resistance to apoptosis and anoikis
Dissemination requires that tumour cells detach from the matrix or cell–cell anchor(s) that control tissue architecture. Under normal circumstances, epithelial cells undergo apoptosis (programmed cell death) when adhesion to the correct substrate is disrupted. Indeed, a specialised form of apoptosis—anoikis—results when normal cells are maintained in suspension; this process is clearly a mechanism designed to protect multicellular organisms from rogue cells establishing themselves outside their correct anatomical location. Metastatic cells therefore must be resistant to anoikis and apoptosis to survive during dissemination and colonisation of ectopic sites. Many studies show that crucial apoptotic modulators are deregulated in metastases. This deregulation is accomplished by various means: activation of survival pathways (eg, PI3 kinase-AKT), upregulation of matrix metalloproteinases (which downregulate death receptors, release growth factors, and condition the extracellular matrix for invasion); overexpression of anti-apoptotic proteins (BCL-2, BCL-XL) or focal adhesion kinase (FAK), and inactivation of p53, among others. The importance of anoikis resistance in metastasis was elegantly shown in experimental studies where a functional screen for suppressors of anoikis identified the neurotrophic receptor TRKB as a key mediator. Rat intestinal epithelial cells are very sensitive to detachment-induced anoikis and are non-tumourigenic, but when transfected with TRKB, they become highly tumourigenic and metastatic via both the lymphatic and haematogenous routes, even destroying bone. TRKB is often overexpressed in human malignancies and is mutated in colon cancer. Its activation also induces vascular endothelial growth factor (VEGF) expression via hypoxia inducible factor (HIF), potentially assisting with establishment and angiogenesis of tumours at secondary sites.
 that control tissue architecture. Under normal circumstances, epithelial cells undergo apoptosis (programmed cell death) when adhesion to the correct substrate is disrupted. Indeed, a specialised form of apoptosis—anoikis—results when normal cells are maintained in suspension; this process is clearly a mechanism designed to protect multicellular organisms from rogue cells establishing themselves outside their correct anatomical location. Metastatic cells therefore must be resistant to anoikis and apoptosis to survive during dissemination and colonisation of ectopic sites. Many studies show that crucial apoptotic modulators are deregulated in metastases. This deregulation is accomplished by various means: activation of survival pathways (eg, PI3 kinase-AKT), upregulation of matrix metalloproteinases (which downregulate death receptors, release growth factors, and condition the extracellular matrix for invasion); overexpression of anti-apoptotic proteins (BCL-2, BCL-XL) or focal adhesion kinase (FAK), and inactivation of p53, among others. The importance of anoikis resistance in metastasis was elegantly shown in experimental studies where a functional screen for suppressors of anoikis identified the neurotrophic receptor TRKB as a key mediator. Rat intestinal epithelial cells are very sensitive to detachment-induced anoikis and are non-tumourigenic, but when transfected with TRKB, they become highly tumourigenic and metastatic via both the lymphatic and haematogenous routes, even destroying bone. TRKB is often overexpressed in human malignancies and is mutated in colon cancer. Its activation also induces vascular endothelial growth factor (VEGF) expression via hypoxia inducible factor (HIF), potentially assisting with establishment and angiogenesis of tumours at secondary sites.")
109
Lancet 2007; 369 Angiogenesis and lymphangiogenesis
That tumour growth and progression is limited before vascularisation of the neoplastic mass is generally accepted.Vascularisation is achieved via neoangiogenesis, co-option of existing blood vessels, vasculogenic mimicry (in which poorly differentiated, highly malignant tumour cells can form a primitive vascular system), or a combination of these processes. Newly formed leaky capillaries can also serve as conduits for disseminating cells. Hypoxia and activated oncogenes, including RAS, EGFR, and HER2/NEU, upregulate angiogenic cytokines (eg, VEGF and interleukin 8) and proteolytic enzymes (eg, matrix metalloproteinases, urokinase plasminogen activator [uPA]) and downregulate inhibitors such as thrombospondin (TSP1) via PI3 kinase and MAP kinase signalling pathways, thus potentiating angiogenesis, tumour growth, and spread. Hypoxia could also directly affect tumour cell motility, invasion, and metastasis, with a key component recently identified as HIF1-regulated lysyl oxidase (LOX). LOX is linked to poor prognosis in several tumour types, including breast and oral cancers. It is thought to regulate FAK activity, cell-matrix adhesion, and motility, potentially creating a niche permissive for metastatic growth at secondary sites. CXCR4, a chemokine implicated in site-selective metastasis, is also upregulated by hypoxia as well as oncogenes such as HER2, MET, and EGFR. Hypoxia regulates many other genes linked to tumour progression, recruits macrophages and other inflammatory cells, and can also contribute to increased genetic instability and resistance to apoptosis. A parallel process—lymphangiogenesis—has been invoked as a potential facilitator of lymphatic metastasis, although functional lymphatic vessels within human tumours are rare and co-option of existing lymphatic vessels could also occur. The major lymphangiogenic cytokines (VEGF-C and VEGF-D) and lymphangiogenesis have been linked to poor prognosis in some cancers, and more specifically with lymph node metastasis. Experimental manipulation of these cytokines modulates lymphatic metastasis in some experimental models. VEGF-A and other signalling systems—eg, angiopoietin:Tie, ephrin:Eph, and PDGF-BB:PDGFR—could also be involved. Recently, so-called zip codes have been identified on the lymphatic endothelium in tumour xenografts which, when blocked, inhibited lymphatic metastasis. Two questions are of particular interest: can the propensity for lymphatic metastasis be predicted from gene expression signatures, as has been claimed for other sites of metastasis? And does lymphatic dissemination predispose to distant metastasis? The role of lymphatic dissemination has recently been reviewed, and that nodal metastases could serve as a bridgehead for further dissemination in certain cancers is clear; however, direct haematogenous dissemination occurs in others. Clearly, more careful kinetic and molecular dissection of cancer spread is required to delineate the importance of tumour cells detected not only in nodes, but also in the blood and bone marrow.
, or a combination of these processes. Newly formed leaky capillaries can also serve as conduits for disseminating cells. Hypoxia and activated oncogenes, including RAS, EGFR, and HER2/NEU, upregulate angiogenic cytokines (eg, VEGF and interleukin 8) and proteolytic enzymes (eg, matrix metalloproteinases, urokinase plasminogen activator [uPA]) and downregulate inhibitors such as thrombospondin (TSP1) via PI3 kinase and MAP kinase signalling pathways, thus potentiating angiogenesis, tumour growth, and spread. Hypoxia could also directly affect tumour cell motility, invasion, and metastasis, with a key component recently identified as HIF1-regulated lysyl oxidase (LOX). LOX is linked to poor prognosis in several tumour types, including breast and oral cancers. It is thought to regulate FAK activity, cell-matrix adhesion, and motility, potentially creating a niche permissive for metastatic growth at secondary sites. CXCR4, a chemokine implicated in site-selective metastasis, is also upregulated by hypoxia as well as oncogenes such as HER2, MET, and EGFR. Hypoxia regulates many other genes linked to tumour progression, recruits macrophages and other inflammatory cells, and can also contribute to increased genetic instability and resistance to apoptosis. A parallel process—lymphangiogenesis—has been invoked as a potential facilitator of lymphatic metastasis, although functional lymphatic vessels within human tumours are rare and co-option of existing lymphatic vessels could also occur. The major lymphangiogenic cytokines (VEGF-C and VEGF-D) and lymphangiogenesis have been linked to poor prognosis in some cancers, and more specifically with lymph node metastasis. Experimental manipulation of these cytokines modulates lymphatic metastasis in some experimental models. VEGF-A and other signalling systems—eg, angiopoietin:Tie, ephrin:Eph, and PDGF-BB:PDGFR—could also be involved. Recently, so-called zip codes have been identified on the lymphatic endothelium in tumour xenografts which, when blocked, inhibited lymphatic metastasis. Two questions are of particular interest: can the propensity for lymphatic metastasis be predicted from gene expression signatures, as has been claimed for other sites of metastasis And does lymphatic dissemination predispose to distant metastasis The role of lymphatic dissemination has recently been reviewed, and that nodal metastases could serve as a bridgehead for further dissemination in certain cancers is clear; however, direct haematogenous dissemination occurs in others. Clearly, more careful kinetic and molecular dissection of cancer spread is required to delineate the importance of tumour cells detected not only in nodes, but also in the blood and bone marrow.")
110
Lancet 2007; 369 Dissemination and colonisation of secondary sites Several million cells per gram of tumour can be shed daily into the lymphatic system or bloodstream. The fate of bloodborne tumour cells is somewhat controversial and experimental evidence contradictory. In some models, most circulating cells die, whereas in others, most survive and extravasate. Insufficient data exist to quantify the fraction of shed tumour cells that successfully seed secondary tissues, especially in human cancers. Nevertheless, all studies show that most cells entering the vasculature fail to form macroscopic foci at distant sites. What, then, is required of a cell to successfully colonise another tissue? These abilities must co-exist within a single cell since metastases are mainly clonal. To accomplish this process, tumour cells use a variety of motility mechanisms, chemokine gradients, and proteinases (eg, matrix metalloproteinases, cathepsins, uPA, etc) to enter and exit the circulation. Many of the processes are also activated by endothelial cells during angiogenesis. Interestingly, Friedl and colleagues recently showed that the migration of tumour cells through collagen matrices still occurs in the presence of broad-spectrum proteinase inhibitor cocktails. Additionally, the intuitive assumption that these enzymes are derived from tumour cells has been challenged by the finding that most are produced by stromal cells. Ultimately, metastatic cells must lodge at secondary sites and re-establish adhesive connections. During haematogenous dissemination, the transit time is only seconds, thus cells are unlikely to shut down transcriptional expression of adhesion molecules as they depart the primary tumour and re-express them when they arrive at secondary sites. Cells could use alternative adhesion molecules or might selectively alter adhesion by post-translational modification of already expressed proteins, glycoproteins, lectins, or other molecules. Metastatic cancer cells must also evade immune effectors or co-opt immune/inflammatory cells to assist them in completing subsequent steps of the metastatic cascade, and they must resist hydrostatic sheer forces (ie, turbulence within vessels). Susceptibility to such stresses can vary widely and could contribute stochastically to metastatic inefficiency. Nonetheless, a successful metastatic cell must overcome whatever challenges to its survival are mounted.
 to enter and exit the circulation. Many of the processes are also activated by endothelial cells during angiogenesis. Interestingly, Friedl and colleagues recently showed that the migration of tumour cells through collagen matrices still occurs in the presence of broad-spectrum proteinase inhibitor cocktails. Additionally, the intuitive assumption that these enzymes are derived from tumour cells has been challenged by the finding that most are produced by stromal cells. Ultimately, metastatic cells must lodge at secondary sites and re-establish adhesive connections. During haematogenous dissemination, the transit time is only seconds, thus cells are unlikely to shut down transcriptional expression of adhesion molecules as they depart the primary tumour and re-express them when they arrive at secondary sites. Cells could use alternative adhesion molecules or might selectively alter adhesion by post-translational modification of already expressed proteins, glycoproteins, lectins, or other molecules. Metastatic cancer cells must also evade immune effectors or co-opt immune/inflammatory cells to assist them in completing subsequent steps of the metastatic cascade, and they must resist hydrostatic sheer forces (ie, turbulence within vessels). Susceptibility to such stresses can vary widely and could contribute stochastically to metastatic inefficiency. Nonetheless, a successful metastatic cell must overcome whatever challenges to its survival are mounted.")
111
TUMOREN - Definities - Ontstaan & evolutie Oorzaken
Precancereuze condities Metastasering - Verdediging - Morfologische diagnose (labotechnieken)
")
112
1) door tumor-suppressor genen
VERDEDIGING Het lichaam kan zichzelf tegen tumoren verdedigen op 3 manieren : 1) door tumor-suppressor genen 2) het immuunsysteem 3) door inflammatie
 door tumor-suppressor genen. 2) het immuunsysteem. 3) door inflammatie.")
Verwante presentaties







![Deltion College Engels C1 Gesprekken voeren [Edu/002]/ subvaardigheid lezen thema: Order, order…. can-do : kan een bijeenkomst voorzitten © Anne Beeker.](/8/2048322/big_thumb.jpg "Deltion College Engels C1 Gesprekken voeren [Edu/002]/ subvaardigheid lezen thema: Order, order…. can-do : kan een bijeenkomst voorzitten © Anne Beeker.>")













