Download de presentatie
1
Klonering in S. cerevisiae en andere gisten en fungi
Saccharomyces cerevisiae : “budding yeast” groeiende uitstulping (“bud”) : afsplitsing van nieuwe cel 16 chromosomen weinig introns groei en ontwikkeling mogelijk als haploïden en als diploïden Schizosaccharomyces pombe : “fission yeast” celdeling : splitsing 3 chromosomen meer introns meer gelijkend op hogere eukaryoten dan S. cerevisiae Methylotrofe gist : Pichia pastoris : hoge expressierendementen Primrose & Twyman : hoofdstuk 11
 : afsplitsing van nieuwe cel. 16 chromosomen. weinig introns. groei en ontwikkeling mogelijk als haploïden en als diploïden. Schizosaccharomyces pombe : fission yeast celdeling : splitsing. 3 chromosomen. meer introns. meer gelijkend op hogere eukaryoten dan S. cerevisiae. Methylotrofe gist : Pichia pastoris : hoge expressierendementen. Primrose & Twyman : hoofdstuk 11.")
2
Klonering in S. cerevisiae
Achtergrond Organisme bij uitstek – Saccharomyces cerevisiae (bakkersgist/biergist) Grootte: 12 Mb (excl. enkele repeats, o.a. rRNA); haploide set chromosomen : 16 Eencellige eukaryoot : doel studie : inzicht in eukaryotische processen recombinante eiwitexpressie /secretie- en fermentatiemogelijkheden geen pyrogene toxines, wel glycosylaties, relatief grote opbrengst, relatief zuiver gemakkelijker te kweken dan dierlijke cellen mogelijkheid klonering grote DNA’s (YAC’s) veel auxotrofe en andere genetische merkers
 Grootte: 12 Mb (excl. enkele repeats, o.a. rRNA); haploide set chromosomen : 16. Eencellige eukaryoot : doel studie : inzicht in eukaryotische processen. recombinante eiwitexpressie /secretie- en. fermentatiemogelijkheden. geen pyrogene toxines, wel glycosylaties, relatief grote opbrengst, relatief zuiver. gemakkelijker te kweken dan dierlijke cellen. mogelijkheid klonering grote DNA’s (YAC’s) veel auxotrofe en andere genetische merkers.")
3
* N Yeast : ribosomale RNAs
Open-leesraam (ORF) analyse van chromosoom XIV (linkerarm) * N Yeast : ribosomale RNAs Alle nucleair-gecodeerde ribosomale RNAs van de Saccharomyces cerevisiae zijn gegroepeerd in één gebied op chromosoom XII. Enkele daarvan zijn in sequentie gebracht. Aan de rechterzijde bevinden zich verschillende kopijen van een 3,6 kb herhaling (precieze aantal in het genoom tot minstens 1997 onbekend), waarin het ASP3 gen en een 5S rRNA gen.
 analyse van chromosoom XIV (linkerarm) * N. Yeast : ribosomale RNAs. Alle nucleair-gecodeerde ribosomale RNAs van de Saccharomyces cerevisiae zijn gegroepeerd in één gebied op chromosoom XII. Enkele daarvan zijn in sequentie gebracht. Aan de rechterzijde bevinden zich verschillende kopijen van een 3,6 kb herhaling (precieze aantal in het genoom tot minstens 1997 onbekend), waarin het ASP3 gen en een 5S rRNA gen.")
4
* Noot : diploïde en haploïde gistcellen
centromeersequenties - chromosomen *

5
Klonering in S. cerevisiae
Transformatie niet “natuurlijk” transformeerbaar (vroeger) aanmaak van sferoplasten/protoplasten (d.i. cellen zonder celwand – enzymatische behandeling) + permeabilisatie met polyethyleenglycol, behandeling met CaCl2 + toevoeging DNA; regeneratie celwand in medium met 3% agar nieuwere methoden: gebruik van lithiumacetaat, PEG, ssDNA (voorbehandeling) ; enzymatische verwijdering niet nodig alternatieven : elektroporatie, conjugatie naar protoplasten geen bruikbare gistvirussen – geen transductie (infectie)/virale vectoren
 aanmaak van sferoplasten/protoplasten (d.i. cellen zonder celwand – enzymatische behandeling) + permeabilisatie met polyethyleenglycol, behandeling met CaCl2 + toevoeging DNA; regeneratie celwand in medium met 3% agar. nieuwere methoden: gebruik van lithiumacetaat, PEG, ssDNA (voorbehandeling) ; enzymatische verwijdering niet nodig. alternatieven : elektroporatie, conjugatie naar protoplasten. geen bruikbare gistvirussen – geen transductie (infectie)/virale vectoren.")
6
Klonering in S. cerevisiae
Vectoren gistreplicon of integratie in een chromosoom of ander replicerend element ; vanuit historisch oogpunt van vectorontwikkeling, was er ook meestal een E. coli plasmide-ori voorzien ; waarom een E. coli plasmide-ori ? : - isolatie van intact plasmide DNA uit S. cerevisiae is moeilijk - bereiding van zuivere en grote hoeveelheden plasmide DNA werd mogelijk na een E. coli transformatie met een (klein, ruw) DNA extract uit gist (dus, E. coli als tussenstap) selectiemerkers? 1) gist is weinig of niet gevoelig voor antibiotica (uitzondering: geneticine G418) geneticine G418 derivaat van neomycine – toxisch substraat voor aminoglycoside fosfotransferasen – prokaryotische enzymen – te voorzien van gistexpressiesignalen bleek actief in gist als resistentiemerker
 DNA extract uit gist (dus, E. coli als tussenstap) selectiemerkers 1) gist is weinig of niet gevoelig voor antibiotica (uitzondering: geneticine G418) geneticine G418. derivaat van neomycine – toxisch. substraat voor aminoglycoside fosfotransferasen – prokaryotische enzymen – te voorzien van gistexpressiesignalen. bleek actief in gist als resistentiemerker.")
7
LEU2 - vereist voor biosynthese van leucine
2) maar meestal werden biosynthetische genen gebruikt – auxotrofe merkers Werkwijze ? Transformeren naar een stam met mutatie in een biosynthetisch gen de stam is afhankelijk van het voedingssupplement Plasmide draagt het wild-type gen na transformatie complementatie van de groeideficiëntie URA3 gen - vereist voor biosynthese van uracil (in feite ribo-uridylaat : rUMP) LEU vereist voor biosynthese van leucine HIS vereist voor biosynthese van histidine Voorbeelden van auxotrofe selectiemerkers en bijhorende biochemische component gist E. coli LEU2 leuB b-isopropylmalaat dehydrogenase URA3 pyrF orotidine-5’-fosfaat decarboxylase HIS3 hisB imidazol glycerolfosfaat dehydratase TRP5 trpAB tryptofaan synthase ARG4 argH argininosuccinaat lyase Terminologie : auxotroof = mutant, niet in staat een bepaalde verbinding nodig voor zijn groei zelf aan te maken, maar groeit wel als die toegevoegd wordt aan het cultuurmedium. prototroof = in staat te groeien op een gedefinieerd minimaal medium, dat idealiter slechts één C-bron en anorganische verbindingen bevat. niet verwarren met : autotroof = haalt zijn C uit CO2 (energie en reducerende kracht uit fotosynthese of chemosynthese) heterotroof = haalt C uit organische verbindingen (en kan ze mineraliseren tot CO2 en H2O)
 maar meestal werden biosynthetische genen gebruikt – auxotrofe merkers. Werkwijze Transformeren naar een stam met mutatie in een biosynthetisch gen. de stam is afhankelijk van het voedingssupplement. Plasmide draagt het wild-type gen. na transformatie complementatie van de groeideficiëntie. URA3 gen - vereist voor biosynthese van uracil (in feite ribo-uridylaat : rUMP) LEU2 - vereist voor biosynthese van leucine. HIS3 - vereist voor biosynthese van histidine. Voorbeelden van auxotrofe selectiemerkers en bijhorende biochemische component. gist E. coli. LEU2 leuB b-isopropylmalaat dehydrogenase. URA3 pyrF orotidine-5’-fosfaat decarboxylase. HIS3 hisB imidazol glycerolfosfaat dehydratase. TRP5 trpAB tryptofaan synthase. ARG4 argH argininosuccinaat lyase. Terminologie : auxotroof = mutant, niet in staat. een bepaalde verbinding nodig. voor zijn groei zelf aan te maken, maar groeit wel als die toegevoegd. wordt aan het cultuurmedium. prototroof = in staat te groeien op een gedefinieerd minimaal medium, dat idealiter slechts één C-bron. en anorganische verbindingen bevat. niet verwarren met : autotroof = haalt zijn C uit CO2 (energie en reducerende kracht uit fotosynthese of chemosynthese) heterotroof = haalt C uit organische verbindingen (en kan ze mineraliseren tot CO2 en H2O)")
8
URA3 – ura3p : systematische naam YEL021W
Beschrijving : orotidine-5‘-fosfaat (OMP) decarboxylase : katalyseert de zesde enzymatische stap in de de novo biosynthese van pyrimidines, door omzetting van OMP tot uridine monofosfaat (UMP) 5-FOA = 5-fluoro-orotic acid (na inbouw) 5-fluoro-uracil Toxic URA3 : is ook een tegenselecteerbare merker : aanwezigheid van URA3-gen is letaal op een medium met 5-FOA
 decarboxylase : katalyseert de zesde. enzymatische stap in de de novo biosynthese van pyrimidines, door omzetting van OMP. tot uridine monofosfaat (UMP) 5-FOA = 5-fluoro-orotic acid. (na inbouw) 5-fluoro-uracil. Toxic. URA3 : is ook een tegenselecteerbare merker : aanwezigheid van URA3-gen is letaal op een medium met 5-FOA.")
9
Kan een gist DNA fragment een auxotrofe E. coli mutant complementeren?
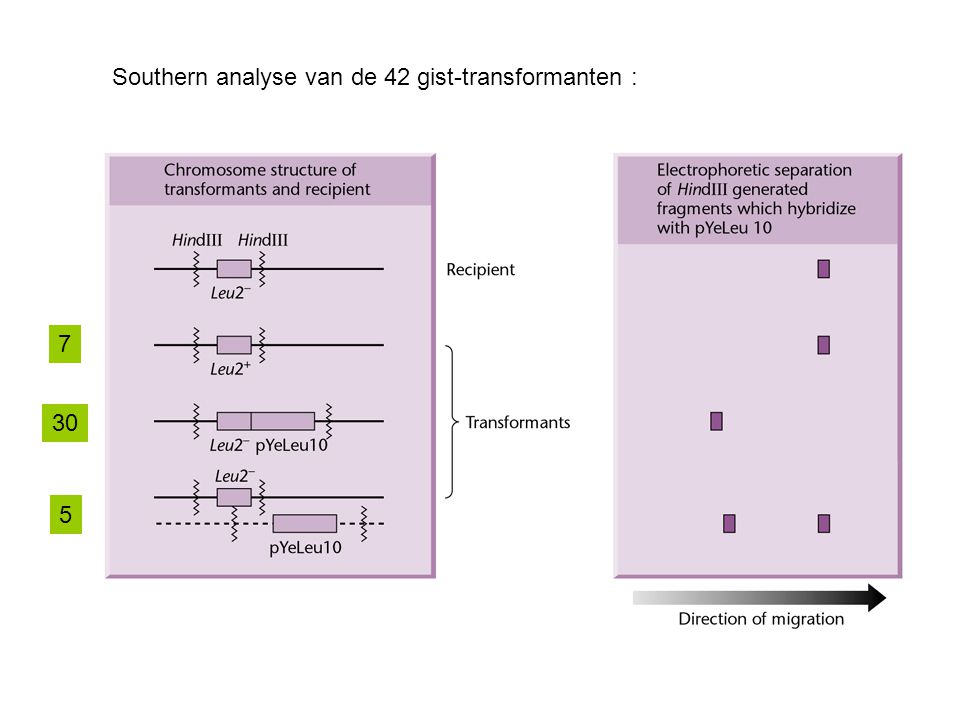
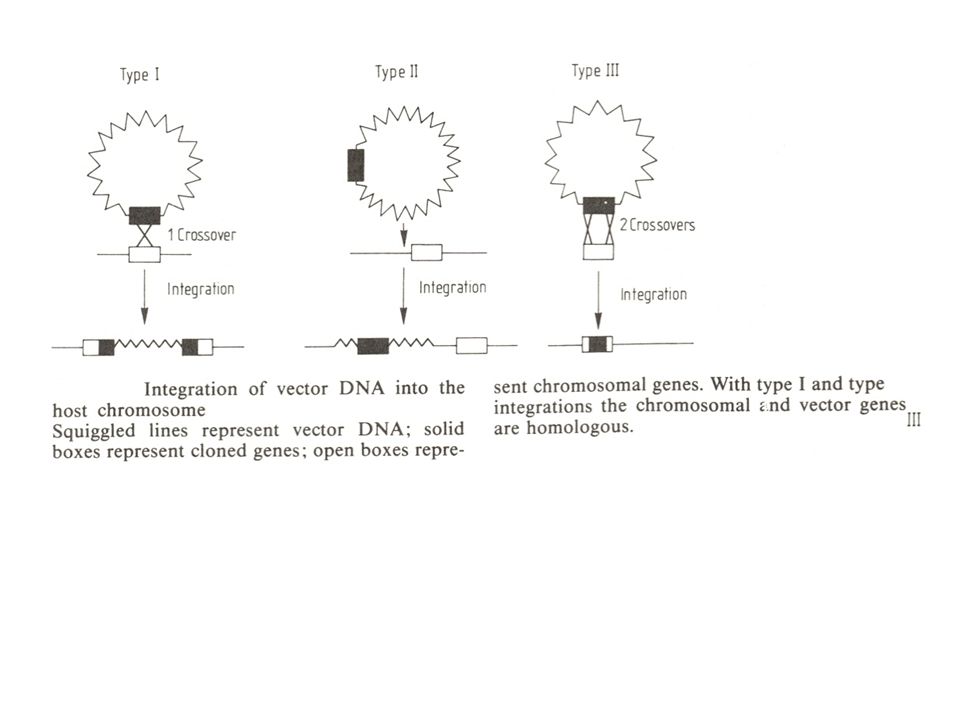
=> test : transformatie van een E. coli leuB stam met restrictiefragmenten van gist DNA => leidde tot identificatie van het gist LEU2 gen Vervolgens omgekeerd : Kan dit gekloneerde LEU2 plasmide DNA een LEU2- mutant complementeren? => test : import in gistcellen 42 transformanten werden bekomen (analyse door Southern hybridisatie) 35 bevatten "pBR322-DNA" (m.a.w. vectorsequenties) (wees op integratie door enkelvoudige recombinatie) 30 op de homologe plaats (type I integratie) 5 op een andere plaats (type II integratie) 7 via allelomwisseling (type III integratie : dubbele homologe recombinatie)
 35 bevatten pBR322-DNA (m.a.w. vectorsequenties) (wees op integratie door enkelvoudige recombinatie) 30 op de homologe plaats (type I integratie) 5 op een andere plaats (type II integratie) 7 via allelomwisseling (type III integratie : dubbele homologe recombinatie)")
10
Southern analyse van de 42 gist-transformanten :
7 30 5
12
2 strategieën voor gerichte integratie :
B. 2 strategieën voor gerichte integratie : lineaire (homologe) uiteinden zijn zeer recombinogeen 1. linearisatie in het doelwitgen en inbouw van de ganse vector 2. voorafgaande inbouw van de merker in het doelwitgen, en excisie van het merkerfragment met doelwitsequenties aan de uiteinden Telkens een intact merkergen, met uiteinden, homoloog aan het doelgebied in het waardcel DNA. Wil men hierbij nog een extra, niet-selecteerbaar gen meebrengen, kan dit uiteraard naast het merkergen (op de vector bij 1. of op het fragment bij 2.) liggen.
 uiteinden zijn zeer recombinogeen. 1. linearisatie in het doelwitgen en inbouw van de. ganse vector. 2. voorafgaande inbouw van de merker in het. doelwitgen, en excisie van het merkerfragment. met doelwitsequenties aan de uiteinden. Telkens een intact merkergen, met uiteinden, homoloog aan het doelgebied in het waardcel DNA. Wil men hierbij nog een extra, niet-selecteerbaar gen meebrengen, kan dit uiteraard naast het merkergen (op de vector bij 1. of op het fragment bij 2.) liggen.")
13
Vectortypes yeast integrating plasmid (YIp) yeast replicating plasmids (YRp) yeast episomal plasmids (YEp) yeast centromere plasmids (YCp) yeast artificial chromosomes (YAC) Primrose: Tabel 11.1
 yeast artificial chromosomes (YAC) Primrose: Tabel")
14
yeast integrating plasmid (YIp)
Vb. YIP5 Antibioticumselectiemerkers voor E. coli: ApR, TcR – pendelen! E. coli ori ColE1 (van pBR322) – pendelen! URA3 - gistselectiemerker (ev. seq voor homologe recombinatie of alternatief : een repetitieve sequentie) “kloneervehikel” voor stabiele introductie van constructen in het gistchromosoom behoud door integratie in genoom (onder selectiedruk) replicatie met het genoom, en zelfde kopijenaantal
 – pendelen! URA3 - gistselectiemerker. (ev. seq voor homologe recombinatie. of alternatief : een repetitieve sequentie) kloneervehikel voor stabiele introductie. van constructen in het gistchromosoom. behoud door integratie in genoom (onder selectiedruk) replicatie met het genoom, en zelfde kopijenaantal.")
15
yeast integrating plasmid (YIp)
Alleluitwisseling met YIp-vectoren uitvissen van mutante genen (allelen) uit het genoom integratie duplicatie van het gen excisie op flankerende knipplaatsen en transformatie naar E. coli leidt tot 50% wt en 50% mutant allel (door linearizatie, efficiëntere integratie – lineaire uiteinden dirigeren de insertieplaats)
 uit het genoom. integratie duplicatie van het gen. excisie op flankerende knipplaatsen en transformatie. naar E. coli leidt tot 50% wt en 50% mutant. allel. (door linearizatie, efficiëntere integratie – lineaire uiteinden dirigeren de insertieplaats)")
16
Integratieve vector : voor stabiele expressie in Pichia pastoris
Zeocine selectie is mogelijk zowel in bacteriën (E. coli), gist als dierlijke cellen. ARG4 mogelijk als (extra) auxotrofe merker ; hier mogelijks in te bouwen in het AOX1 doelwitgen (AOX1 = aldehyde oxidase 1) TT : transcriptieterminator Zeocine behoort tot de bleomycine/phleomycine familie van antibiotica die binden aan DNA (intercaleren) en het afbreken. Resistentie steunt op het Sh ble genproduct (van Streptoalloteichus hindustanus), dat bindt aan het antibioticum en aldus de binding aan DNA verhindert.
, gist als dierlijke cellen. ARG4 mogelijk als (extra) auxotrofe merker ; hier mogelijks in te bouwen in het AOX1 doelwitgen. (AOX1 = aldehyde oxidase 1) TT : transcriptieterminator. Zeocine behoort tot de bleomycine/phleomycine familie van antibiotica die binden aan DNA (intercaleren) en het afbreken. Resistentie steunt op het Sh ble genproduct (van Streptoalloteichus hindustanus), dat bindt aan het antibioticum en aldus de binding aan DNA verhindert.")
17
Recombinogene klonering in gist.
Homologe recombinatie is zeer efficient in S. cerevisiae (en ook behoorlijk in andere fungi) en ds-uiteinden zijn het beste substraat. Dit kan uitgebuit worden door fragmenten met corresponderende uiteinden in vivo te laten recombineren (in dit voorbeeld in een gistvector in de cel) en aldus DNA segmenten uit te wisselen (bvb. een merker, een genconstruct, enz.). REMI : restriction enzyme mediated integration Lineaire DNA fragmenten, bvb. BamHI fragmenten, die samen met het BamHI restrictieënzym aan een transformatiemengsel toegevoegd worden, zullen (bij voorkeur) integreren in een genomische BamHI plaats. De transformatie-efficiëntie is tevens sterk verhoogd. Deze techniek is ook toegepast in andere fungi.
 en. ds-uiteinden zijn het beste substraat. Dit kan uitgebuit worden door fragmenten met. corresponderende uiteinden in vivo te laten. recombineren (in dit voorbeeld in een gistvector in. de cel) en aldus DNA segmenten uit te wisselen. (bvb. een merker, een genconstruct, enz.). REMI : restriction enzyme mediated integration. Lineaire DNA fragmenten, bvb. BamHI fragmenten, die samen met het BamHI restrictieënzym aan een. transformatiemengsel toegevoegd worden, zullen. (bij voorkeur) integreren in een genomische BamHI. plaats. De transformatie-efficiëntie is tevens sterk. verhoogd. Deze techniek is ook toegepast in andere fungi.")
18
yeast replicating plasmid (YRp)
Vb. YRP17 E. coli elementen (fig/tabel) intermediair kopijaantal - meer transformanten ARS = ‘Autonomous Replicating Sequences’ - replicatie in gist gemeenschappelijk motief 5’WTTTATRTTTW3’ URA3, TRP1 selectiemerkers nadeel : mindere stabiliteit en neiging tot segregatie met de oudercel oudercel - ondanks de selectiedruk
 intermediair kopijaantal - meer transformanten. ARS = ‘Autonomous Replicating. Sequences’ - replicatie in gist. gemeenschappelijk motief. 5’WTTTATRTTTW3’ URA3, TRP1 selectiemerkers. nadeel : mindere stabiliteit en neiging. tot segregatie met de oudercel. oudercel - ondanks de selectiedruk.")
19
(A) Structuur van ARS1, een typisch autonoom replicerend element(ARS) dat werkt als een ori in Saccharomyces cerevisiae. De relatieve posities van de functionele sequenties A, B1, B2 en B3 zijn aangeduid. (B) Opensmelten van de DNA helix gebeurt in subdomein B2,geïnduceerd door aanhechting van het ARS bindingsproteïne 1 (ABFI1) aan subdomein B3. De eiwitten van het “origin replication complex” zijn permanent gebonden aan subdomeinen A en B1. *
 Opensmelten van de DNA helix gebeurt in subdomein B2,geïnduceerd door aanhechting van het ARS bindingsproteïne 1 (ABFI1) aan subdomein B3. De eiwitten van het origin replication complex zijn permanent gebonden aan subdomeinen A en B1. *")
20
yeast replicating plasmid (YRp)
Toepassing: alleluitwisseling YRP17 7002 bp ORI : Col E1 ori voor E. coli Ap : bla ; ampicilline resistentiegen Tc : tet ; tetracycline resistentiegen

21
Alleluitwisseling zonder integratie !
‘Genconversie’-mechanisme ! (allel dient als “matrijs” voor aanmaak van nieuwe seq.) Vb. Isolatie van een genomisch allel van de MAT locus : ‘uitvissen’ van de allelsequentie uit het genoom linearisatie / behoud van de flankerende sequenties dubbele cross-over Gelineariseerd DNA kan enkel blijven bestaan in gist na circularisatie of integratie => hercircularisatie na recombinatie en genduplicatie - “pendeling” naar E. coli en analyse van de mutanten
 Vb. Isolatie van een genomisch allel van de MAT. locus : ‘uitvissen’ van de allelsequentie uit het genoom. linearisatie / behoud van de flankerende sequenties. dubbele cross-over. Gelineariseerd DNA kan enkel blijven bestaan in. gist na circularisatie of integratie. => hercircularisatie na recombinatie. en genduplicatie. - pendeling naar E. coli en analyse van de mutanten.")
22
yeast episomal plasmid (YEp)
Vb. YEp24 Hoog kopijenaantal ( per cel) E. coli plasmide-elementen URA3 selectiemerker in gist Replicatie in gist door replicatie-elementen van het circulaire 2µ plasmide 2µ plasmide – vrij algemeen voorkomend in S. cerevisiae ~ 6300 bp (naamgeving : 2m, 2-m, 2mu, 2-mu, 2mm, ...) replicon en stabiliteitslocus vereist voor vectorconstructie (ORI + REP3) niet erg stabiel, maar toch beter dan de YRp vectoren.
 E. coli plasmide-elementen. URA3 selectiemerker in gist. Replicatie in gist door replicatie-elementen van het circulaire 2µ plasmide. 2µ plasmide – vrij algemeen voorkomend in S. cerevisiae. ~ 6300 bp (naamgeving : 2m, 2-m, 2mu, 2-mu, 2mm, ...) replicon en stabiliteitslocus vereist voor vectorconstructie (ORI + REP3) niet erg stabiel, maar toch beter dan de YRp vectoren.")
23
De twee vormen van het 2m plasmide : het centrale gedeelte is de omgekeerde herhaling
(inverted repeat) ORI en REP3 zijn vereist in cis. REP1 en REP2 kunnen in trans voorzien worden : => in CIR+ cellen volstaat 1 à 1,5 kb 2m DNA om een repliceerbaar YEp plasmide te maken. CIR+ cellen bevatten 2m plasmide, CIRo cellen bevatten geen 2m.
 ORI en REP3 zijn vereist in cis. REP1 en REP2 kunnen in trans voorzien. worden : => in CIR+ cellen volstaat 1 à 1,5 kb. 2m DNA om een repliceerbaar. YEp plasmide te maken. CIR+ cellen bevatten 2m plasmide, CIRo cellen bevatten geen 2m.")
24
yeast episomal plasmid (YEp)
deel van het circulair 2m plasmide YEp24 7769 bp

25
yeast centromere plasmids (YCp)
Stabilisatie van een YRp door een CEN (centromeer) sequentie : ongeveer 120bp in budding yeast vormt een “minichromosoom” : - stabiel in mitose (>10 kb) (selectie op stabilisatie) - segregeert mendeliaans in meïose - 1 kopij per cel
 sequentie : ongeveer 120bp in budding yeast. vormt een minichromosoom : - stabiel in mitose (>10 kb) (selectie op stabilisatie) - segregeert mendeliaans in. meïose. - 1 kopij per cel.")
26
yeast artificial chromosome (YAC)
lineair DNA, met centromeersequentie, telomeersequenties, ARS, selectiemerkers (cfr. klonering van grote DNAs) YAC-vectoren : ‘yeast’ artificieel chromosoom (gist) (1987) "Klassieke" YACs : lineair DNA : architectuur zoals gist chromosoom in 1987 werd een kloneringsprocedure uitgewerkt : YAC vectoren Latere ontwikkelingen : circulaire (of gecirculariseerde) YAC’s TAR : (transformation-associated recombination) Zie hoofdstuk 14 : vectoren voor klonering van grotere DNA’s
 YAC-vectoren : ‘yeast’ artificieel chromosoom (gist) (1987) Klassieke YACs : lineair DNA : architectuur zoals gist chromosoom. in 1987 werd een kloneringsprocedure uitgewerkt : YAC vectoren. Latere ontwikkelingen : circulaire (of gecirculariseerde) YAC’s. TAR : (transformation-associated recombination) Zie hoofdstuk 14 : vectoren voor klonering van grotere DNA’s.")
27
Na bepaling van de totale DNA-sequentie van het gistgenoom :
systematische gendisruptie en aanleggen van ORF-gedeleteerde stammen (‘deletant strains’) (onderzoek naar het aantal 'echte' genen en hun functie) Doel : - genfuncties bestuderen door aanleggen van disruptanten en fenotypische analyse van de mutanten onderscheid tussen essentiële en niet-essentiële genen bepalen collecties van mutanten: haploïde van "mating " a en a type homozygote diploïden voor de niet-essentiële genen heterozygote diploïden voor de essentiële genen Strategie: PCR-gebaseerde deletiecassette : KanMX-module: aminoglycosidefosfotransferase o.i.v. "gist"-transcriptiesequenties selectie op geneticine G418 Studie van essentiële genen (16-18%) via temperatuursensitieve mutanten (bvb. geninactivatie leidt tot afsterven - aanmaak van temperatuursgevoelige allelen : puntmutaties (‘error-prone PCR’) die invloed hebben op de eiwitstructuur/stabiliteit (vb. cs/ts-mutanten = ‘cold sensitive’/’temperature sensitive’ mutanten) : mutante eiwitten destabiliseren bij 16°C.
 (onderzoek naar het aantal echte genen en hun functie) Doel : - genfuncties bestuderen door aanleggen van disruptanten en fenotypische analyse. van de mutanten. onderscheid tussen essentiële en niet-essentiële genen bepalen. collecties van mutanten: haploïde van mating a en a type. homozygote diploïden voor de niet-essentiële genen. heterozygote diploïden voor de essentiële genen. Strategie: PCR-gebaseerde deletiecassette : KanMX-module: aminoglycosidefosfotransferase o.i.v. gist -transcriptiesequenties. selectie op geneticine G418. Studie van essentiële genen (16-18%) via temperatuursensitieve mutanten. (bvb. geninactivatie leidt tot afsterven - aanmaak van temperatuursgevoelige allelen : puntmutaties (‘error-prone PCR’) die invloed hebben op de eiwitstructuur/stabiliteit. (vb. cs/ts-mutanten = ‘cold sensitive’/’temperature sensitive’ mutanten) : mutante eiwitten. destabiliseren bij 16°C.")
28
Gendisruptie – aanleggen van gen-gedeleteerde stammen (‘deletant strains’) van gist
(PCR-gebaseerd) Constructie
 Constructie.")
29
Gendisruptie – aanleggen van gen-gedeleteerde stammen (‘deletant strains’) - GIST
Verificatie

30
Strategie voor aanbrengen van grote
deleties in het S. cerevisiae genoom. Split-marker vectoren : stuffer-1 stuffer-2 U R A - uitsplitsen met NotI - transformatie van gistcellen genoom U R A 3 - 3-dubbele recombinatie in gist - selectie naar URA3 (doel : vermijden van (selectie van) type II integratie)
 type II integratie)")
31
* N Addendum Humane genen (en bijhorende kunnen complementeren.
genproducten) die gist-mutanten kunnen complementeren. (update mei 1994) * N
 die gist-mutanten. kunnen complementeren. (update mei 1994) * N.")


>")

















