Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Plaatsgerichte mutagenese
Primrose & Twyman Principles of Gene Manipulation and Genomics, 7e editie (2006), Hoofdstuk 8 - doel: enkelvoudige of enkele posities gericht wijzigen in een DNA fragment (kloon) (enkelvoudige, gerichte wijzigingen in een genoom vergt bijzondere technieken: LATER) - 'reverse genetics' : klassieke genetica : willekeurig mutaties invoeren om fenotype te creëren => plaats van mutatie zoeken (en karteren) 'reverse' genetica : mutatie invoeren op welbepaalde plaats => mogelijk effect op fenotype (functie) bekijken - drie basisbenaderingen: (1) cassettemutagenese (2) primerverlenging (3) PCR
, Hoofdstuk 8. - doel: enkelvoudige of enkele posities gericht wijzigen in een DNA fragment (kloon) (enkelvoudige, gerichte wijzigingen in een genoom vergt bijzondere technieken: LATER) - reverse genetics : klassieke genetica : willekeurig mutaties invoeren om fenotype te creëren. => plaats van mutatie zoeken (en karteren) reverse genetica : mutatie invoeren op welbepaalde plaats. => mogelijk effect op fenotype (functie) bekijken. - drie basisbenaderingen: (1) cassettemutagenese (2) primerverlenging (3) PCR.")
2
(klassieke) genetica causaliteit DNA fenotype ‘reverse genetics’
 genetica causaliteit DNA fenotype ‘reverse genetics’")
3
Relatie tussen mutagenese-efficiëntie en het aantal kloons, nodig om de
aanwezigheid van de gewenste mutant te verzekeren, met 90% probabiliteit

4
Cassettemutagenese - excisie van een DNA fragment (restrictieknipplaatsen !) + vervanging door een chemisch-gesynthetiseerd fragment (2 of meer oligonucleotiden) - VOOR - hoge efficiëntie - multipele uitwisseling mogelijk, inclusief degeneraties - TEGEN - vereist flankerende "unieke" knipplaatsen - synthesecapaciteit moet voldoende hoog zijn - gebruik overlappende sets oligonucleotiden mogelijk (cfr. chemische synthese) - bij complexe degeneraties: 2de streng aanmaken door fill-in (rekening houden met hybridisatiekinetiek !) - gebruik van inosine mogelijk om degeneraties te beperken - ook deleties en grote inserties zijn mogelijk
 - VOOR - hoge efficiëntie. - multipele uitwisseling mogelijk, inclusief degeneraties. - TEGEN - vereist flankerende unieke knipplaatsen. - synthesecapaciteit moet voldoende hoog zijn. - gebruik overlappende sets oligonucleotiden mogelijk (cfr. chemische synthese) - bij complexe degeneraties: 2de streng aanmaken door fill-in. (rekening houden met hybridisatiekinetiek !) - gebruik van inosine mogelijk om degeneraties te beperken. - ook deleties en grote inserties zijn mogelijk.")
5
mismatch posities Uitwisseling van restrictiefragmenten

6
gebruik van inosine op gerandomiseerde posities

7
Mismatch primerverlenging ('primer extension')
=> oligonucleotide-gestuurd, mismatch-afhankelijk - vereist enkelstreng matrijs (eventueel partiëel enkelstrengig) - M13- of fasmide kloon - aanmaak van gapped-duplex - vereist mismatch oligonucleotide (chemische synthese) - belang van positie van mismatch in oligonucleotide op matrijs - 3'-effect (repair) => keuze polymerase ! - 5'-effect (displacement) => keuze polymerase ! - mogelijkheden: puntmutatie, multipele puntmutaties, insertie, deletie ('sticky feet'-mutagenese) - efficiëntie is (zeer) laag tenzij bijzondere maatregelen genomen worden waarom ? => afhankelijk van diverse factoren - transformatie door heteroduplex + oorspronkelijke matrijs (ss) => de oorspronkelijke (ss) matrijs geeft alleen niet-mutante transformanten - herstelmechanismen in E.coli (GATC !) => gebruik van mutL, mutS, mutH stammen
 - M13- of fasmide kloon. - aanmaak van gapped-duplex. - vereist mismatch oligonucleotide (chemische synthese) - belang van positie van mismatch in oligonucleotide op matrijs effect (repair) => keuze polymerase ! - 5 -effect (displacement) => keuze polymerase ! - mogelijkheden: puntmutatie, multipele puntmutaties, insertie, deletie. ( sticky feet -mutagenese) - efficiëntie is (zeer) laag tenzij bijzondere maatregelen genomen worden. waarom => afhankelijk van diverse factoren. - transformatie door heteroduplex + oorspronkelijke matrijs (ss) => de oorspronkelijke (ss) matrijs geeft alleen niet-mutante transformanten. - herstelmechanismen in E.coli (GATC !) => gebruik van mutL, mutS, mutH stammen.")
8
Mutagenese door mismatch primer verlenging op een enkelstrengige
matrijs T C

9
Enkelvoudige of meervoudige puntmutaties, insertiemutations en deletiemutaties
zijn mogelijk. Hoofdbekommernis is de efficiëntie van screening.

10
Theoretisch concept, geeft 50% wild-type en 50 mutante nakomelingen.
maar : ook de enkelstrengige moleculen transformeren de waardcel, zodat meer wild-type kloons worden gevormd ; bovendien : herstelmechanismen favoriseren de originele sequentie.

11
Rol van GATC sequenties in mismatch repair : strengselectiviteit

12
Selectie van de mutante streng : belangrijkste voorbeelden
(vaak ook gemodificeerde oligonucleotiden of modificaties in de mismatch primer mogelijk, bvb. met inosine) - methode van Eckstein : - in vitro fixering van de mutatie - gebruik van fosforothioaat nucleotiden (aS-dCTP) - kenmerken van restrictieënzymen tegenover S-modificatie - methode van Kunkel : - in vivo selectie van de mutant - "doping“ van matrijsstreng met uridylaat : 1 à 2 % U waar T thuishoort - dubbelmutant : dut (dUTPase) ung (uracil-N-glycosidase) - transformatie van de mismatch dubbelstreng naar een wt-E.coli
 - methode van Eckstein : - in vitro fixering van de mutatie. - gebruik van fosforothioaat nucleotiden (aS-dCTP) - kenmerken van restrictieënzymen tegenover S-modificatie. - methode van Kunkel : - in vivo selectie van de mutant. - doping van matrijsstreng met uridylaat : 1 à 2 % U waar T thuishoort. - dubbelmutant : dut (dUTPase) ung (uracil-N-glycosidase) - transformatie van de mismatch dubbelstreng naar een wt-E.coli.")
13
Methode van Eckstein Introduceer een S-gemodificeerde streng
Sommige restrictieënzymen worden geïnhibiteerd door S-modificatie, maar splitsen wel nog de niet-gemodificeerde streng (=> nicking) Exonuclease III verwijdert de mismatch, maar alleen uit de parentale streng
 Exonuclease III verwijdert de mismatch, maar alleen uit de parentale streng.")
14
Structuur van dCTPaS : het Sp isomeer wordt specifiek gebruikt door E
Structuur van dCTPaS : het Sp isomeer wordt specifiek gebruikt door E. coli DNA polymerase I

15
methode van Kunkel voor plaatsgerichte mutagenese
De biochemie achter de methode van Kunkel voor plaatsgerichte mutagenese Metabolisme van dUTP in E. coli Pijlen geven de richting aan van de omzettingen in de algemene pathway eerder dan het evenwicht bij de individuele reacties. Pijlen in vetjes duiden de majeure pathway aan. In de kadertjes staan de namen van de betrokken genen : ung : uracyl-DNA N-glycosylase dut : dUTPase dcd : dCTP deaminase cdd : (deoxy)cytidine deaminase deoA : thymidine phosphorylase (deoxyuridine) dCyd : deoxycytidine ; dUrd : deoxyuridine ; dThd : deoxythymidine dRib-1-P : deoxyribose-1-fosfaat
cytidine deaminase. deoA : thymidine phosphorylase. (deoxyuridine) dCyd : deoxycytidine ; dUrd : deoxyuridine ; dThd : deoxythymidine. dRib-1-P : deoxyribose-1-fosfaat.")
16
Vergelijking tussen strategie
van Eckstein en van Kunkel. Kunkel : => de matrijsstreng wordt gemodificeerd ; => "manipulaties" in vivo Eckstein : => de nieuwe streng wordt gemodificeerd; => manipulaties in vitro

17
De ‘gapped duplex’ methode voor in vitro mutagenese
Omwisseling van waardcel tussen “amber suppressing” en “non-suppressing” fenotype. Selectie door gebruik van de niet-suppresserende waardcel.

18
- gapped duplex methode, met extra (= begeleidende) mismatch tgo de complementaire streng
- transformatie naar een niet-suppressor stam (Su-) - expressie vanaf de parentale streng vereist een amber suppressor - gapped duplex method, met alternerende ambermutaties : - Apam => ApR en CmR => Cmam, en vice versa - transformatie naar een Su- stam - maakt consecutieve mutageneses makkelijker - in vivo selectie met een wijziging in de selectiemerker - toevoeging van een selectieprimer die het bla gen wijzigt - mutante bla gen geeft resistentie tegen ceftazidime + ampicilline - niet-mutante transformanten overleven niet op ceftazidime - T4 polymerase vult de openingen (gaps) nauwkeurig in (geen 5'-3' exo, geen ‘strand displacement’) - er is een goede koppeling tussen de mismatchen m.a.w. geen cross-over : aanwezigheid van beide gaat samen - met EcoK en EcoB herkenningssequenties kan omwisseling van 1 bp de (in vivo) restrictiegevoeligheid omkeren A A C n n n n n n G T G C EcoK T G A n n n n n n n n T G C T EcoB T R A C n n n n n n G T G C T samen
 - expressie vanaf de parentale streng vereist een amber suppressor. - gapped duplex method, met alternerende ambermutaties : - Apam => ApR en CmR => Cmam, en vice versa. - transformatie naar een Su- stam. - maakt consecutieve mutageneses makkelijker. - in vivo selectie met een wijziging in de selectiemerker. - toevoeging van een selectieprimer die het bla gen wijzigt. - mutante bla gen geeft resistentie tegen ceftazidime + ampicilline. - niet-mutante transformanten overleven niet op ceftazidime. - T4 polymerase vult de openingen (gaps) nauwkeurig in. (geen 5 -3 exo, geen ‘strand displacement’) - er is een goede koppeling tussen de mismatchen. m.a.w. geen cross-over : aanwezigheid van beide gaat samen. - met EcoK en EcoB herkenningssequenties kan omwisseling van 1 bp de (in vivo) restrictiegevoeligheid omkeren. A A C n n n n n n G T G C EcoK. T G A n n n n n n n n T G C T EcoB. T R A C n n n n n n G T G C T samen.")
19
“Gapped duplex” methode met fenotypische selectie
Omwisseling tussen twee resistentiegenen, rekening houdend met de suppressie-fenotypes. Volle balk : wild-type gen Open balk : ambermutant

20
Selectie van mutanten door een extra mutatie in
een bla gen zodat ceftazidime resistentie ontstaat. De "begeleidende" mutatie wijzigt het bla gen mutante TEM bla S K G 5´ p AAA TCT GGA GCC TCC AAG GGT GGG TCT CGC 3´ aaa tct gga gcc GGT GAG Cgt ggg tct cgc wild-type TEM bla G E R

21
Ceftazidime Ceftazidime is a third-generation cephalosporin antibiotic. Like other third-generation cephalosporins, it has broad spectrum activity against Gram-positive and Gram-negative bacteria. Unlike most third-generation agents, it is active against Pseudomonas aeruginosa, however, it has weaker activity against Gram-positive microorganisms and is not used for such infections. Ceftazidime is a semisynthetic, broad-spectrum, b-lactam antibiotic for parenteral administration. Ceftazidime is bactericidal in action exerting its effect by inhibition of enzymes responsible for cell-wall synthesis. A wide range of gram-negative organisms is susceptible to ceftazidime in vitro, including strains resistant to gentamycin and other aminoglycosides. In addition, ceftazidime has been shown to be active against gram-positive organisms. It is highly stable to most clinically important b-lactamases, plasmid or chromosomal, which are produced by both gram-negative and gram-positive organisms and, consequently, is active against many strains resistant to ampicillin and other cephalosporins. Ceftazidime has activity against the gram-negative organisms Pseudomonas and Enterobacteriaceae. Its activity against Pseudomonas is a distinguishing feature of ceftazidime among the cephalosporins.

22
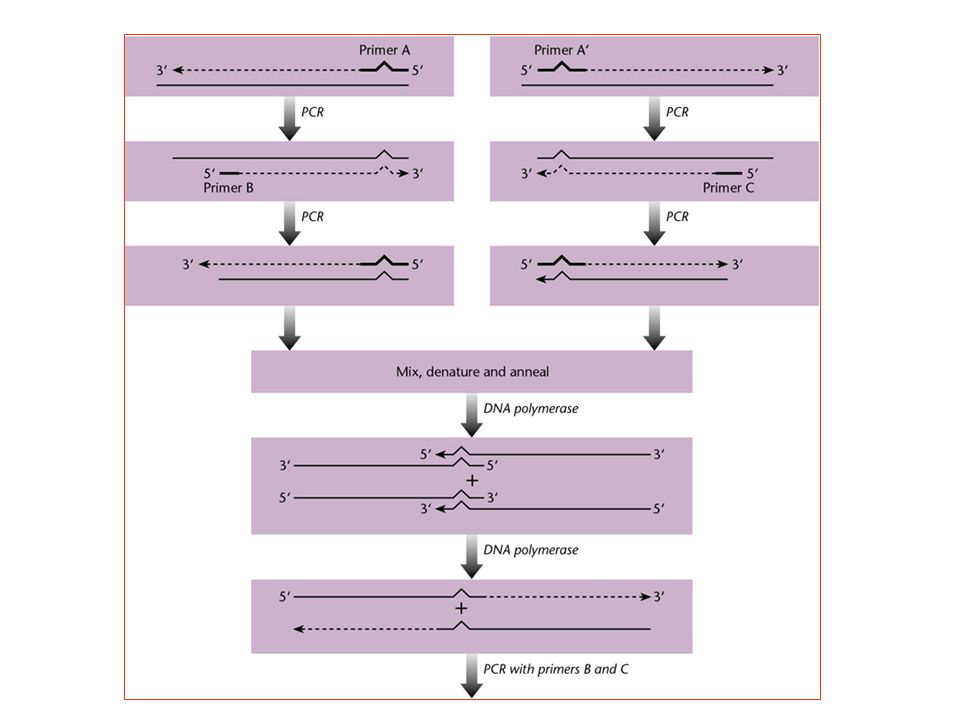
PCR-gebaseerde plaats-specifieke mutagenese
- uitschakeling van de parentale matrijs door amplificatie - mismatch primer op de doelwitpositie => bij “terugpriming” (in 2nd cyclus) wordt de wijziging gefixeerd - uitbreiding van cassettemutagenese, maar de regio’s (fragmenten) kunnen veel groter zijn ; restrictieplaats "in de omgeving" van de mismatch is nodig, maar dat mag niettemin op een redelijke afstand zijn (bvb. 25 of zelfs meer nucleotiden, zolang de primer synthetiseerbaar blijft) - de nood aan restrictieplaats kan omzeild worden door een werkwijze die sterk op SOE lijkt - 'megaprimer' mutagenese: tweestapsprocedure, waarin het vroege ampliconproduct de primer wordt in de daaropvolgende amplificatie - door inverse PCR : indien de vector klein genoeg is - lineair product - circularisatie nodig - een splitsingsplaats is weerom zeer nuttig - primers met staarten maken substantiële inserties mogelijk - “universele” methode met Class-II restrictieplaatsen en enzymen
 wordt de wijziging gefixeerd. - uitbreiding van cassettemutagenese, maar de regio’s (fragmenten) kunnen veel groter zijn ; restrictieplaats in de omgeving van de mismatch is nodig, maar dat mag. niettemin op een redelijke afstand zijn (bvb. 25 of zelfs meer nucleotiden, zolang. de primer synthetiseerbaar blijft) - de nood aan restrictieplaats kan omzeild worden door een werkwijze die sterk op SOE lijkt. - megaprimer mutagenese: tweestapsprocedure, waarin het vroege ampliconproduct. de primer wordt in de daaropvolgende amplificatie. - door inverse PCR : indien de vector klein genoeg is. - lineair product. - circularisatie nodig. - een splitsingsplaats is weerom zeer nuttig. - primers met staarten maken substantiële inserties mogelijk. - universele methode met Class-II restrictieplaatsen en enzymen.")
24
De megaprimer methode De mutante moleculen gevormd in de eerste PCR-rondes fungeren als primers in de latere cycli van de PCR.

25
Aanpassingen van “inverse PCR” :

26
Gebruik van EarI of andere ClassII-S enzymen
EarI : afsplitsing van extra "staarten" C T C T T C (1 / 4) herkenningssequentie van EarI ----CTCTTCn ----GAGAAGnnnn
 herkenningssequentie van EarI. ----CTCTTCn. ----GAGAAGnnnn.")
27
Andere alternatieven : - ExciteTM methode : tegenselectie met DpnI
- GeneTailorTM methode : in vitro C-methylatie en in vivo afbraak van parentale streng De PCR methoden zijn een majeure benadering geworden voor gerichte mutagenese - VOOR - efficientie zo goed als 100% (onder optimale condities) - eenvoud - TEGEN - het product is lineair: inbouw in een vector of circularisatie is nodig om het amplicon te kloneren - risico van extra mutaties (in de rest van het amplicon) => sequencing van het ganse amplicon => keuze van polymerase is belangrijk ... veel andere alternativen, met ondermeer meervoudige gerichte mutaties op een welbepaalde plaats in een koderende sequenties, insertie van ongewone aminozuren, gene shuffling, enz.
 - eenvoud. - TEGEN - het product is lineair: inbouw in een vector of circularisatie is. nodig om het amplicon te kloneren. - risico van extra mutaties (in de rest van het amplicon) => sequencing van het ganse amplicon. => keuze van polymerase is belangrijk. ... veel andere alternativen, met ondermeer meervoudige gerichte mutaties. op een welbepaalde plaats in een koderende sequenties, insertie van. ongewone aminozuren, gene shuffling, enz.")
28
'ExciteTM' -methode Plasmide DNA uit een dam+ stam
Inverse PCR met 5'-gefosforyleerde primers (T4-kinase + ATP) Behandeling met DpnI vernietigt de parentale strengen (GmeATC) (inclusief hybride) Circularisatie met ligase en transformatie van E. coli
 Behandeling met DpnI vernietigt de parentale. strengen (GmeATC) (inclusief hybride) Circularisatie met ligase. en transformatie van E. coli.")
29
'GeneTailor' -methode In vitro methylatie van het plasmide DNA (meC)
Amplificatie via inverse PCR met overlappende 5'-uiteinden (+ één mismatch primer) Transformatie van E. coli wt-stam (mcrBC+) McrBC endonuclease vernietigt de gemethyleerde matrijsstreng De 'repair'-enzymen circulariseren het linaire product.
 Transformatie van E. coli wt-stam (mcrBC+) McrBC endonuclease vernietigt de. gemethyleerde matrijsstreng. De repair -enzymen circulariseren het linaire. product.")
30
Het maken van fusies tussen gen-1 en gen-2

31
Creëren of verwijderen van een restrictieknipplaats als hulp
bij screening naar plaatsspecifieke mutanten Doel : vervangen Trp door Phe Hulp bij screening : MnlI splitsing Mismatch primer : …GCCCTGGGCTTCGGTGGCA…

32
Codewoord tabel voor ‘reverse translation’
Leu = 1T Arg = 3G Ser = 567 any aa

33
Regio-gerichte mutagenese :
Error-prone PCR : verhogen van de foutieve inbouw door Taq polymerase Wijziging van reactiecondities, vooral Mg2+/Mn2+ concentraties en ongelijke concentraties tussen de 4 dNTP's. Mutatiefrequenties (misincorporaties) mogelijke tot 1/300 bp en meer. Suppressie van ambermutaties : SupF (tyrT mutant) bouwt tyrosine in, ongeacht of het oorspronkelijke aminozuur op de amber-mutante positie stond. SupE (glnV mutant) bouwt glutamine in, SupD een serine, SupU tryptofaan, SupP leucine, enz. Men heeft gesleuteld aan de corresponderende genen om dit spectrum te verruimen (tot de meeste van de 20 aminozuren). Inbouw van "onnatuurlijke" aminozuren : amber en opaal codons zijn mogelijke doelwitten. Wijziging van een archaea tRNA zodat UAG of UGA als codon herkend worden. Het bijhorende aminoacyl-tRNA synthetase wordt gemuteerd ("ge-engineered") opdat het een speciaal aminozuur (exclusief !) zou opladen : bvb. fenylalanine- of tyrosinederivaten. m-acetylphenylalanine in de plaats van Lys7 in proteïne Z. m-acetylphenylalanine inde plaats van Arg200 in LamB.
 mogelijke tot 1/300 bp en meer. Suppressie van ambermutaties : SupF (tyrT mutant) bouwt tyrosine in, ongeacht of. het oorspronkelijke aminozuur op de amber-mutante positie stond. SupE (glnV mutant) bouwt glutamine in, SupD een serine, SupU tryptofaan, SupP leucine, enz. Men heeft gesleuteld aan de corresponderende genen om dit. spectrum te verruimen (tot de meeste van de 20 aminozuren). Inbouw van onnatuurlijke aminozuren : amber en opaal codons zijn. mogelijke doelwitten. Wijziging van een archaea tRNA zodat UAG of UGA als codon. herkend worden. Het bijhorende aminoacyl-tRNA synthetase wordt. gemuteerd ( ge-engineered ) opdat het een speciaal aminozuur. (exclusief !) zou opladen : bvb. fenylalanine- of tyrosinederivaten. m-acetylphenylalanine in de plaats van Lys7 in proteïne Z. m-acetylphenylalanine inde plaats van Arg200 in LamB.")
34
Detectie (bevestiging)
- fysisch: ontstaan of verdwijnen van knipplaats (restrictieanalyse) (reverse-translation analyse naar mogelijke manipulatie) - sequencing: extra primer nodig op korte afstand van de mismatch positie - hybridisatie: +/- analyse met de mismatch-primer
 (reverse-translation analyse naar mogelijke manipulatie) - sequencing: extra primer nodig op korte afstand van de mismatch positie. - hybridisatie: +/- analyse met de mismatch-primer.")
Verwante presentaties



















>")