Download de presentatie
De presentatie wordt gedownload. Even geduld aub

2
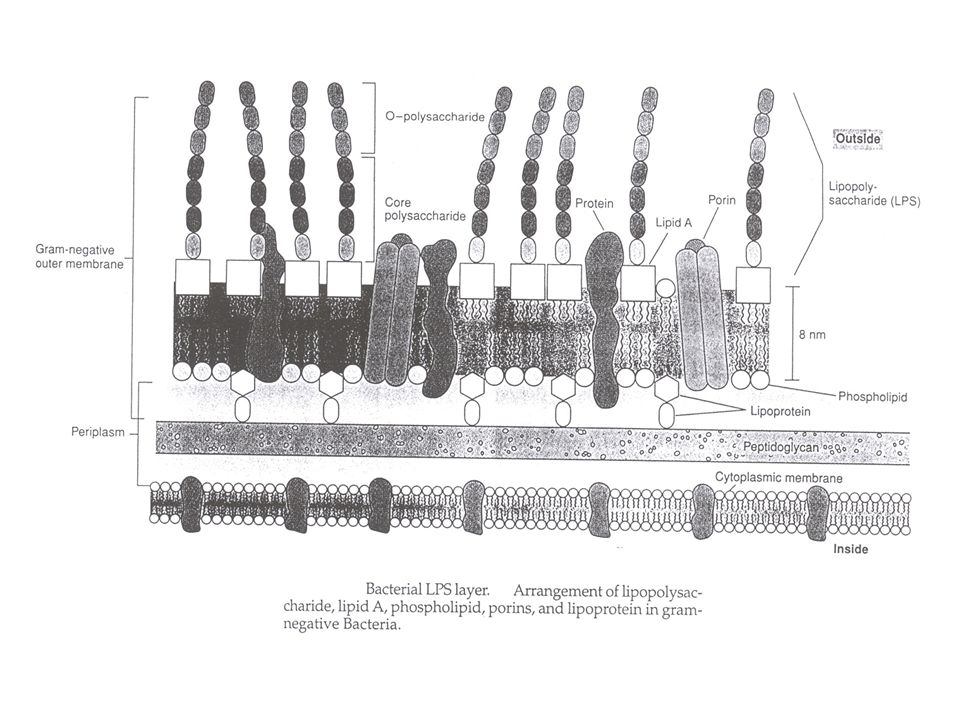
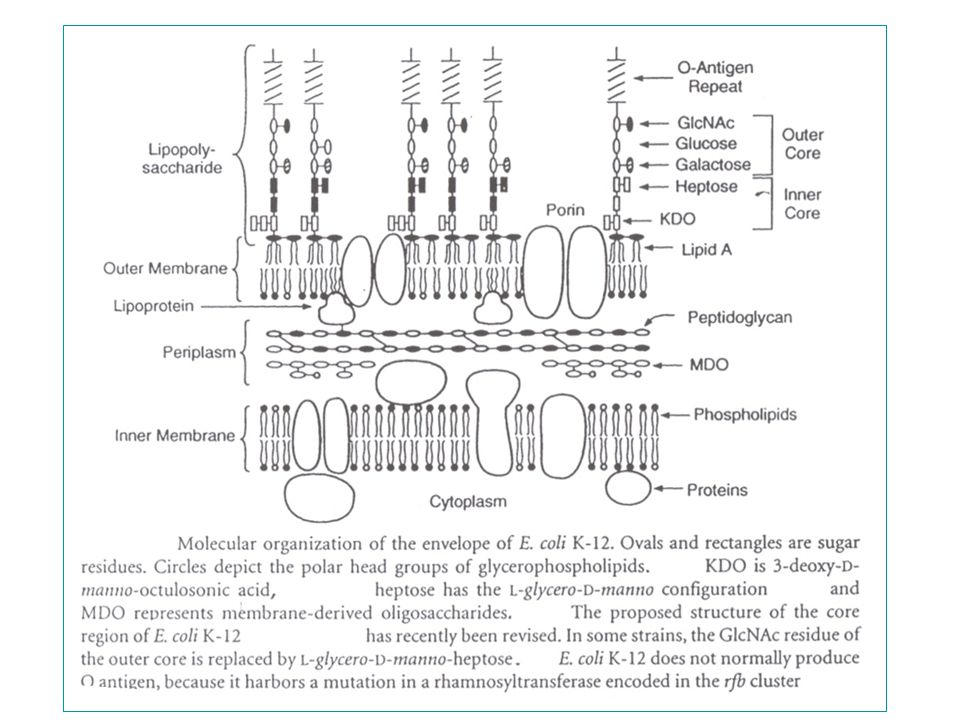
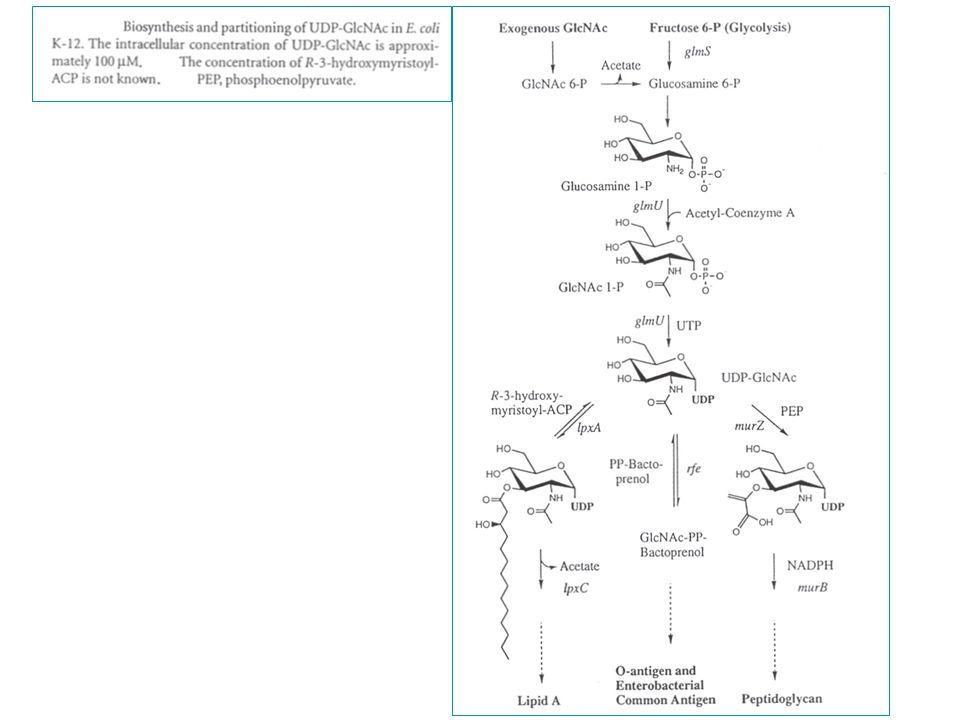
LPS - immunogene glycolipiden in de buitenste membraan van Gram-negatieve bacteriën - vrij complex : minstens 50 genen betrokken bij de assembly => immunogeniciteit (variatie) : O-antigeen => pyrogeniciteit (endotoxine) : lipide A Bestaat uit 3 covalent-verbonden domeinen: 1) lipide A, het endotoxine - is het hydrofobe anker in de buitenste membraan - is bioactieve component verantwoordelijk voor bepaalde pathofysiologische aspecten van Gram-negatieve bacteriële infecties - is endotoxine : => pyrogeniciteit (verwekt koorts in dieren bij injectie) 2) het "core" gebied : een gefosforyleerd, niet-repeterend oligosaccharide - heeft te maken met barrière tegen een aantal antibiotica (zonder core-gebied: hypersensitiviteit aan antibiotica) - inner core (met Kdo en heptose suikers) en outer core 3) O-antigeen polymeer : de determinantenketen (naam: OHNE flagellen) - een immunogene herhaling (tussen 1 en 40) van een oligosaccharide - variabel van stam tot stam - (E. coli K12 : geen O-antigeensynthese door een insertiemutatie in de rfb cluster, nl. een mutatie in het rhamnosyltransferasegen)
 : O-antigeen. => pyrogeniciteit (endotoxine) : lipide A. Bestaat uit 3 covalent-verbonden domeinen: 1) lipide A, het endotoxine. - is het hydrofobe anker in de buitenste membraan. - is bioactieve component verantwoordelijk voor bepaalde pathofysiologische. aspecten van Gram-negatieve bacteriële infecties. - is endotoxine : => pyrogeniciteit (verwekt koorts in dieren bij injectie) 2) het core gebied : een gefosforyleerd, niet-repeterend oligosaccharide. - heeft te maken met barrière tegen een aantal antibiotica. (zonder core-gebied: hypersensitiviteit aan antibiotica) - inner core (met Kdo en heptose suikers) en outer core. 3) O-antigeen polymeer : de determinantenketen (naam: OHNE flagellen) - een immunogene herhaling (tussen 1 en 40) van een oligosaccharide. - variabel van stam tot stam. - (E. coli K12 : geen O-antigeensynthese door een insertiemutatie in de rfb. cluster, nl. een mutatie in het rhamnosyltransferasegen)")
6
- er zijn mutanten van E. coli K12 en andere Gram-negatieve bacteriën die
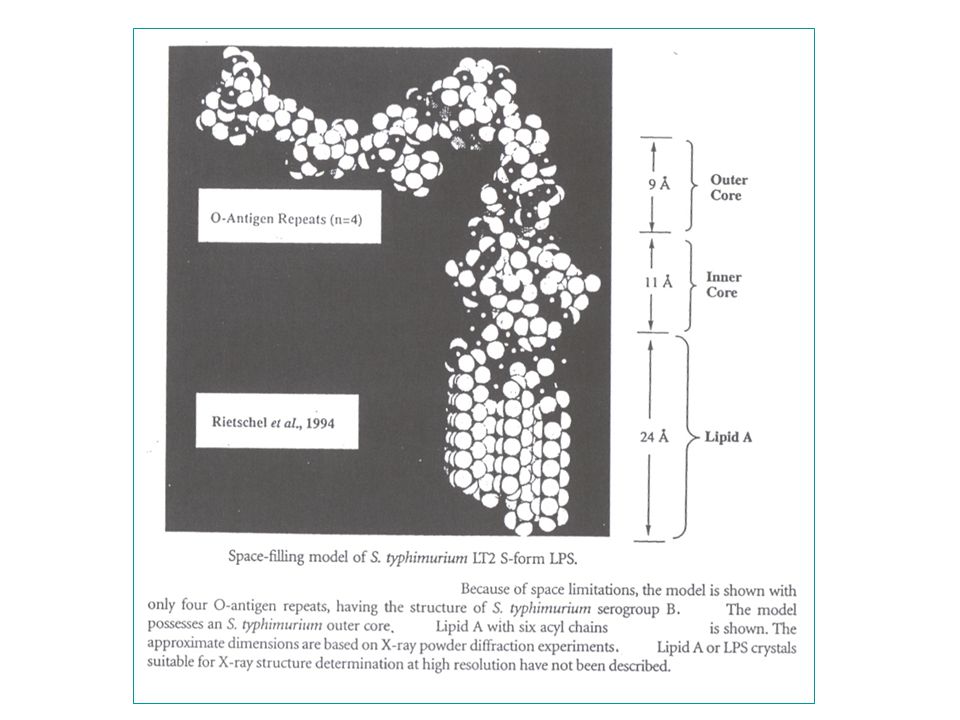
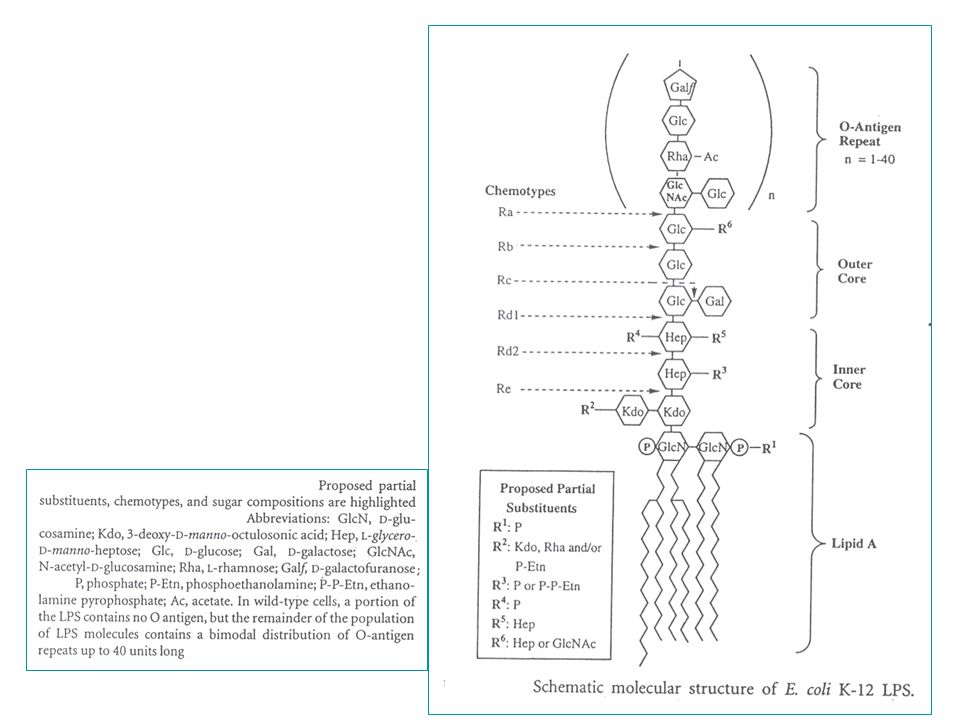
getrunceerd zijn tot in de core : => chemotypes (Ra, Rb, enz.) - de minimale structuur van LPS = Lipide A Kdo eenheden = chemotype Re : 24 chirale centra - centraal in de ontrafeling van de LPS structuur staat de (onverwachte) ontdekking (1983) van lipide X : (2,3 diacylglucosamine-1-fosfaat) een monosaccharide precursor van lipide A die accumuleerde in bepaalde fosfatidylglycerol-deficiënte mutanten van E. coli.
 - de minimale structuur van LPS = Lipide A + 2 Kdo eenheden = chemotype Re : 24 chirale centra. - centraal in de ontrafeling van de LPS structuur staat de (onverwachte) ontdekking (1983) van lipide X : (2,3 diacylglucosamine-1-fosfaat) een monosaccharide precursor. van lipide A die accumuleerde in bepaalde fosfatidylglycerol-deficiënte mutanten. van E. coli.")
9
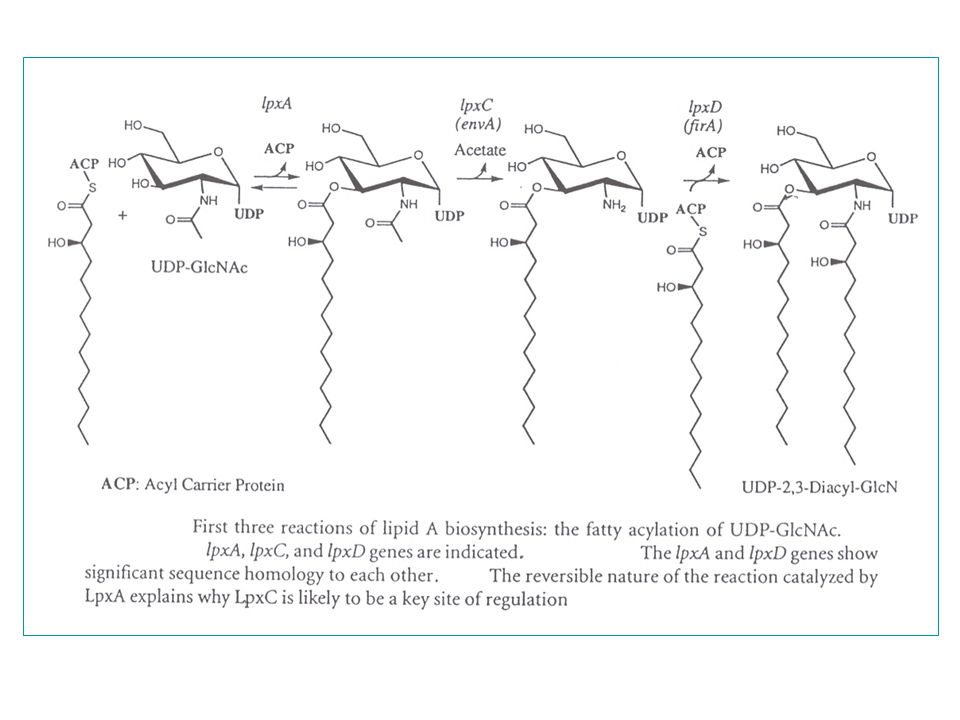
Opbouw (opgehelderd in 1987)
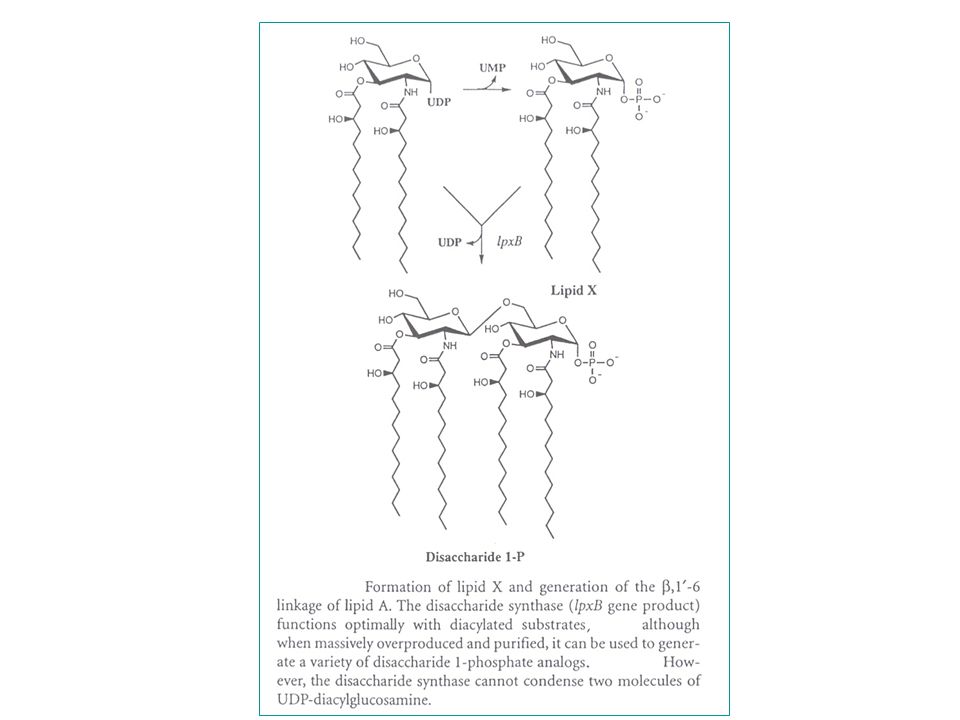
stap 1 : NAG-UDP - (3) acylatie (3-OH-myristinezuur C14 keten) lpxA evenwicht naar links - (2 N) deacetylatie (regulatie! : drijft vorige reactie naar rechts) lpxC - (2 N) acylatie (tweede 3-OH-myristinezuur) lpxD stap 2 : - pyrofosfaatsplitsing (deactivatie) (monosacch-UDP => monosacch-P) (LIPIDE X) - koppeling : monosacch-UDP + monosacch-P => disacch-P b, 1’ lpxB (LIPIDE A) stap 3 : - fosforylatie op C4' : => P-disacch-P (kinase) substraat ATP - aanhechting: Kdo (1) (Kdo-transferase) kdtA - bifunctioneel enzym : lipide IVA en Kdo-lipide IVA zijn substraat - Kdo-CMP : zeer labiele precursor (halfwaardetijd in water +/- 10 min) - aanhechting: Kdo (2) (idem) - andere Gram-negatieve bacteriën = andere aantallen Kdo (zie verder) acylketens (laureaat, myristaat) htrB en msbB Verdere stappen: - rfa-genengroep voor verdere afwerking van de core - rfb-genengroep voor opbouw van het O-antigeen polymeer
 acylatie (3-OH-myristinezuur C14 keten) lpxA evenwicht naar links. - (2 N) deacetylatie (regulatie! : drijft vorige reactie naar rechts) lpxC. - (2 N) acylatie (tweede 3-OH-myristinezuur) lpxD. stap 2 : - pyrofosfaatsplitsing (deactivatie) (monosacch-UDP => monosacch-P) (LIPIDE X) - koppeling : monosacch-UDP + monosacch-P => disacch-P. b, 1’-6 lpxB (LIPIDE A) stap 3 : - fosforylatie op C4 : => P-disacch-P (kinase) substraat ATP. - aanhechting: Kdo (1) (Kdo-transferase) kdtA. - bifunctioneel enzym : lipide IVA en Kdo-lipide IVA zijn substraat. - Kdo-CMP : zeer labiele precursor (halfwaardetijd in water +/- 10 min) - aanhechting: Kdo (2) (idem) - andere Gram-negatieve bacteriën = andere aantallen Kdo (zie verder) acylketens (laureaat, myristaat) htrB en msbB. Verdere stappen: - rfa-genengroep voor verdere afwerking van de core. - rfb-genengroep voor opbouw van het O-antigeen polymeer.")
11
- Pyrofosfatase om 1 geactiveerde voorloper om te zetten tot het vereiste substraat
voor het disaccharide synthase (LpxB) (LpxB is niet in staat 2 geactiveerdevoorlopers te gebruiken) - Lipide X synthese is 500x verhoogd in conditionele lpxB mutanten. - Lipide X (en allicht ook alle andere monosaccharide-precursoren van lipide A) vormen met water micellen, terwijl lipide A disacchariden assembleren tot dubbellagen (bilayers). Dit zou kunnen verklaren waarom alle enzymen vanaf het disaccharide synthase (LpxB) de aanwezigheid van detergent vereisen voor enzymatische activiteit (in vitro), terwijl de enzymactiviteiten in de stappen ervoor dit niet vereisen.
 (LpxB is niet in staat 2 geactiveerdevoorlopers te gebruiken) - Lipide X synthese is 500x verhoogd in conditionele lpxB mutanten. - Lipide X (en allicht ook alle andere monosaccharide-precursoren van lipide A) vormen met water micellen, terwijl lipide A disacchariden assembleren. tot dubbellagen (bilayers). Dit zou kunnen verklaren waarom alle. enzymen vanaf het disaccharide synthase (LpxB) de aanwezigheid. van detergent vereisen voor enzymatische activiteit (in vitro), terwijl. de enzymactiviteiten in de stappen ervoor dit niet vereisen.")
12
- 4'-kinase absoluut vereist.
transfer van g-fosfaat van ATP op de C4' positie (ook nodig om lipideA substructuren als endotoxine antagonist of mimetica te laten fungeren) - Hoe gevonden? - op 21 min van de genoomkaart van E.coli kende men twee genen (msbA (een translocator) en kdsB (zie verder)) die een operon vormden met een derde ORF (orfE met onbekende functie) - aanwezig in de Kohara library van het E. coli genoom (geordende l-bank) : als een 20 kb fragment : => klonering van orfE in een recombinant expressiesysteem (met T7-promotor) => dit gaf een 2000x hogere 4' kinase-activiteit in gewassen membranen van E.coli cellen => fosforyleert efficient een aantal modelsubstraten/analogen => het orfE werd daarom lpxK genoemd
 - Hoe gevonden - op 21 min van de genoomkaart van E.coli kende men twee genen (msbA (een. translocator) en kdsB (zie verder)) die een operon vormden met een derde ORF. (orfE met onbekende functie) - aanwezig in de Kohara library van het E. coli genoom (geordende l-bank) : als een 20 kb fragment : => klonering van orfE in een recombinant expressiesysteem (met. T7-promotor) => dit gaf een 2000x hogere 4 kinase-activiteit in gewassen. membranen van E.coli cellen. => fosforyleert efficient een aantal modelsubstraten/analogen. => het orfE werd daarom lpxK genoemd.")
14
Kdo complexiteit onder Gram-negatieven : WaaA
(Kdo : afkorting van de oorspronkelijke triviale benaming 2-keto, 3-deoxy-octulonaat) Kdo = 3-deoxy D-manno oct-2-ulosonaat => KdtA : Kdo transferase Kds : Kdo synthase (A en B) - KdtA (kdtA) van E. coli : bifunctioneel enzyme : hecht 2 Kdo's consecutief aan. - gseA is het overeenkomstige gen van Chlamydia trachomatis : het transfereert 3 Kdo's - nadat het gen bij meer en meer microörganismen gevonden werd het kdtA-gen een algemene benaming gegeven: waaA gen (cfr. WaaC en WaaF voor heptosyltransferase I en II)
 Kdo = 3-deoxy D-manno oct-2-ulosonaat. => KdtA : Kdo transferase Kds : Kdo synthase (A en B) - KdtA (kdtA) van E. coli : bifunctioneel enzyme : hecht 2 Kdo s consecutief aan. - gseA is het overeenkomstige gen van Chlamydia trachomatis : het transfereert 3 Kdo s. - nadat het gen bij meer en meer microörganismen gevonden werd het kdtA-gen een. algemene benaming gegeven: waaA gen. (cfr. WaaC en WaaF voor heptosyltransferase I en II)")
15
Complementatie-experimenten met een kdtA::kan mutatie in E. coli:
(kan : = cassette met kanamycineresistentiegen) Bij mutanten met dit allel (op het chromosoom) is celgroei en Kdo-transfer afhankelijk van de aanwezigheid van een plasmide met een intacte kopij van kdtA. Als dit plasmide is een ts-replicon draagt, wordt de groei geïnhibeerd na enkele uren bij 44°C (en de Kdo-transferactiviteit wordt ondetecteerbaar) (Terzelfdertijd treedt een massieve accumulatie van lipideIVA op.) Experiment met C. trachomatis gseA : het hierboven vermelde E. coli mutant allel (kdtA::kan) was complementeerbaar met een plasmide met gseA. LPS van dergelijke cellen reageert fel met antilichamen die tegen een genus-specifiek epitoop van Chlamydia gericht zijn (en waartegen parentaal E. coli LPS niet reageert, m.a.w. dat allicht gericht is tegen een structuur die met het derde Kdo te maken heeft.) Legionella pneumophila : heeft 2 Kdo's complementatie van het E. coli kdtA::kan allel is mogelijk => dit is toch wel verrassend aangezien de lipide A struktuur bij Legionella zeer verschillend is : b, 1-6 verbinding tussen 2 di-aminoglucosen : één op positie 2, en één op positie 3, m.a.w. amide-koppelingen van de acylgroepen, die C14-C22 zijn ; bis-gefosforyleerd)
 Bij mutanten met dit allel (op het chromosoom) is celgroei en Kdo-transfer afhankelijk van de. aanwezigheid van een plasmide met een intacte kopij van kdtA. Als dit plasmide is een ts-replicon draagt, wordt de groei geïnhibeerd na enkele uren bij 44°C. (en de Kdo-transferactiviteit wordt ondetecteerbaar) (Terzelfdertijd treedt een massieve accumulatie van lipideIVA op.) Experiment met C. trachomatis gseA : het hierboven vermelde E. coli mutant allel (kdtA::kan) was complementeerbaar met een plasmide met gseA. LPS van dergelijke cellen reageert fel. met antilichamen die tegen een genus-specifiek epitoop van Chlamydia gericht zijn (en. waartegen parentaal E. coli LPS niet reageert, m.a.w. dat allicht gericht is tegen een structuur. die met het derde Kdo te maken heeft.) Legionella pneumophila : heeft 2 Kdo s. complementatie van het E. coli kdtA::kan allel is mogelijk. => dit is toch wel verrassend aangezien de lipide A struktuur bij Legionella zeer verschillend. is : b, 1-6 verbinding tussen 2 di-aminoglucosen : één op positie 2, en één op. positie 3, m.a.w. amide-koppelingen van de acylgroepen, die C14-C22 zijn ; bis-gefosforyleerd)")
16
gluco-geconfigureerd aminosuiker en een fosfaat op 4' van het
=> dit suggereert de hypothese dat alle Kdo-transferasen een gemeenschappelijk acceptormotief voor de catalytische activiteit hebben, dat gelocaliseerd is in het carbohydraat gedeelte: minstens een b, 1’-6 disaccharide met een gluco-geconfigureerd aminosuiker en een fosfaat op 4' van het niet-reducerende suikerdeel (O-antigeen bij Legionella is ook raar : residus van een derivaat van 5,7 diamino – 3,5,7,9 tetradeoxy-nonulosonaat) H. influenzae WaaA : kan (biochemisch) slechts 1 Kdo transfereren ==> er is in de LPS structuur ook maar 1 Kdo aanwezig. complementeert de E. coli knockout mutant (van de Re chemotype stam) NIET => dit lijkt logisch ... maarrrr…. Bordetella pertussis ("whooping cough") WaaA transfereert 1 Kdo complementeert de E.coli mutant WEL !!!! ==> daarom: terugkijken naar H. influenzae:
 H. influenzae. WaaA : kan (biochemisch) slechts 1 Kdo transfereren. ==> er is in de LPS structuur ook maar 1 Kdo aanwezig. complementeert de E. coli knockout mutant (van de Re chemotype stam) NIET. => dit lijkt logisch ... maarrrr…. Bordetella pertussis ( whooping cough ) WaaA transfereert 1 Kdo. complementeert de E.coli mutant WEL !!!! ==> daarom: terugkijken naar H. influenzae:")
17
==> daarom : terugkijken naar H. influenzae :
1 Kdo : er is een fosfaat op positie C-4, of op positie C-5 (nb. kan ook fosfo-ethanolamine zijn op C-4, maar hier niet verder van belang) het waaA werd gekloneerd : zoals reeds gezegd geeft het geen complementatie van de E.coli knockout mutant, maar er bleek WEL complementatie mogelijk indien samen met het waaA gen ook co-expressie werd uitgevoerd met het kdkA gen van H.influenzae : dit kdkA gen is een LPS-specifiek Kdo-kinasegen, en het KdkA genproduct fosforyleert exclusief op C-4 => dus mogelijks volstaat ook 1 Kdo in E.coli als Kdo positie C-4 gefosforyleerd is. (let wel : complementatie zegt niet a priori hoe het product (LPS) eruit ziet.) waaA van Chlamydophila trachomatis, pneumoniae en psittaci met het C.psittaci gen : hoofdzakelijk LPS met 4 Kdo's (Kdo 2->4 Kdo 2->4 Kdo, met vooraan een 2->8 vierde Kdo) Complementatie van het E. coli allel. Acinetobacter baumanii (en haemolyticus) : in vitro : 2 Kdo's op een synthetische E.coli lipide A voorloper, maar 3 Kdo's indien de hoeveelheid substraat zeer beperkt was. nb. expressie van deze enzymen doet men vooral (bijna uitsluitend) in de grampositieve bacterie Corynebacterium glutamicum waarom?
 het waaA werd gekloneerd : zoals reeds gezegd geeft het geen complementatie. van de E.coli knockout mutant, maar er bleek WEL complementatie mogelijk indien. samen met het waaA gen ook co-expressie werd uitgevoerd met het kdkA gen van. H.influenzae : dit kdkA gen is een LPS-specifiek Kdo-kinasegen, en het KdkA. genproduct fosforyleert exclusief op C-4. => dus mogelijks volstaat ook 1 Kdo in E.coli als Kdo positie C-4 gefosforyleerd is. (let wel : complementatie zegt niet a priori hoe het product (LPS) eruit ziet.) waaA van Chlamydophila trachomatis, pneumoniae en psittaci. met het C.psittaci gen : hoofdzakelijk LPS met 4 Kdo s (Kdo 2->4 Kdo 2->4 Kdo, met vooraan een 2->8 vierde Kdo) Complementatie van het E. coli allel. Acinetobacter baumanii (en haemolyticus) : in vitro : 2 Kdo s op een synthetische E.coli. lipide A voorloper, maar 3 Kdo s indien de hoeveelheid substraat zeer beperkt was. nb. expressie van deze enzymen doet men vooral (bijna uitsluitend) in de grampositieve bacterie Corynebacterium glutamicum. waarom")
18
Synthese van Kdo ribulose-5-P (ketose) => arabinose-5-P (aldose) + PEP => bovenaan COOH, 2-keto, 3-deoxy, OH-4 tot 7 in manno vorm (links-links-rechts-rechts) circularisatie : pyranose : OH-6 naar keto-2 gen kdsA (Kdo synthase gen) : cytoplasmatisch (nb. Kdo transferase is membranair) Vervolgens defosforylatie van C-8 fosfaat, en het product wordt door kdsB-genproduct KdsB met CTP geactiveerd tot Kdo-CMP. activatie met CMP : er is geen fosfaatgroep op C-1 : vandaar geen CDP maar CMP in de structuur (CTP => CMP + PPi) CMP-Kdo is ongebruikelijk labiel : moet ter plaatse aangemaakt worden uit CTP en Kdo, met enzyme in overmaat (halfwaarde CMP-Kdo : ongeveer 10 minuten in vitro) Goed bestudeerd: inhibitoren gekend (cfr. figuur)
 circularisatie : pyranose : OH-6 naar keto-2. gen kdsA (Kdo synthase gen) : cytoplasmatisch (nb. Kdo transferase is membranair) Vervolgens defosforylatie van C-8 fosfaat, en het product wordt door kdsB-genproduct KdsB. met CTP geactiveerd tot Kdo-CMP. activatie met CMP : er is geen fosfaatgroep op C-1 : vandaar geen CDP maar. CMP in de structuur (CTP => CMP + PPi) CMP-Kdo is ongebruikelijk labiel : moet ter plaatse aangemaakt worden uit CTP en Kdo, met enzyme in overmaat (halfwaarde CMP-Kdo : ongeveer 10 minuten in vitro) Goed bestudeerd: inhibitoren gekend (cfr. figuur)")
19
Genetische organisatie
3 genenclusters : lpx eerste stappen lpxA,B,C,D : aparte promotoren nb. rfe ligt elders op de kaart (buiten deze clusters) rfa core suikers 3 operons (2 convergente + kdtA) rfb O antigeen suikers (*) alleen Kdo-transferase en het veronderstelde O-antigeen ligase hebben membraan-overspannende domeinen. De andere enzymen hebben geen hydrofobe gebieden en zijn dus allicht cytoplasmatische of perifere membraanproteïnen. Het veronderstelde O-antigeen ligase is deleteerbaar : dus niet-essentieel voor afwerking van de core.
 rfa core suikers 3 operons (2 convergente + kdtA) rfb O antigeen suikers. (*) alleen Kdo-transferase en het veronderstelde O-antigeen ligase hebben. membraan-overspannende domeinen. De andere enzymen hebben. geen hydrofobe gebieden en zijn dus allicht cytoplasmatische of. perifere membraanproteïnen. Het veronderstelde O-antigeen ligase is deleteerbaar : dus niet-essentieel. voor afwerking van de core.")
20
Verderop in de inner core vinden we heptose :
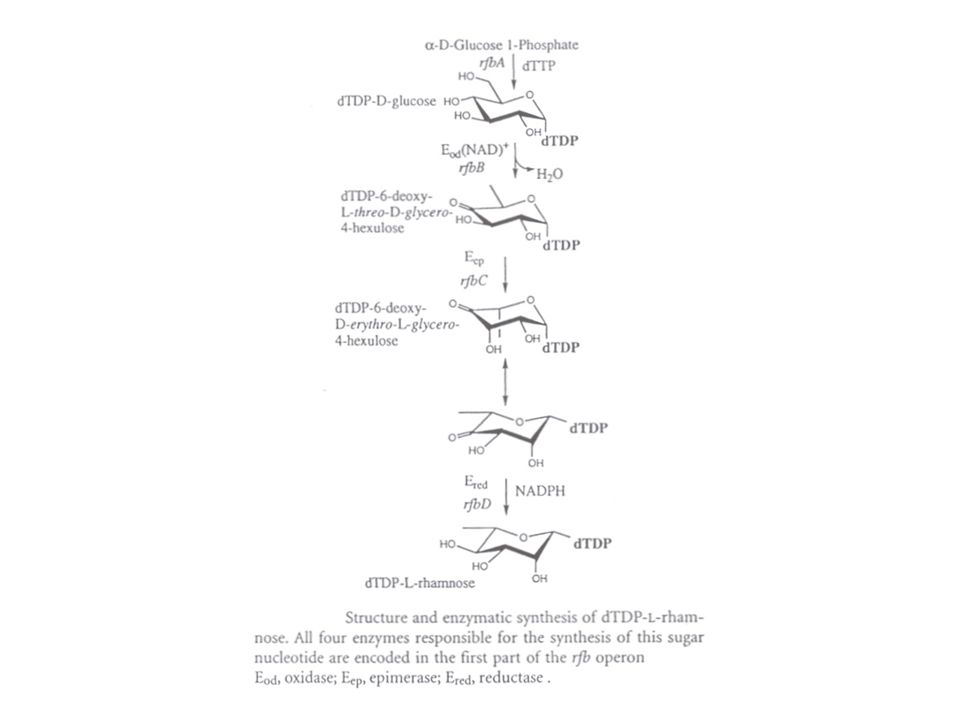
aanmaak : vanuit sedoheptulose-7-fosfaat : eerste stap : isomerase tot pyranosestructuur (vgl. Kdo) verder: mutase, pyrofosforylase, epimerase (na activatie) activatie met ATP tot ADP-suiker : ADP-L-glycero-D-manno-heptose (na activatie wordt één van de OH groepen omgelegd door het epimerase RfaD) tal van decoraties ("versierselen") : fosfo-ethanolamine, enz. verderop (outer core, O-antigen) : tal van bijzondere suikers: (dehydratatie, epimerisatie, reductie) GDP-mannose ==== GDP-L-fucose TDP-glucose ==== TDP-L-rhamnose CDP-glucose ==== CDP-abequose, CDP-paratose, CDP-tyvelose vorming van deoxysuikers vanaf verschillende geactiveerde hexosen (deoxy, dideoxy) polymerisatie met transferasen die in de membraan zitten : een soort "heen en weer drijven" tot de geschikte donor gevonden wordt. (cfr. teichoïnezuursynthese)
 verder: mutase, pyrofosforylase, epimerase (na activatie) activatie met ATP tot ADP-suiker : ADP-L-glycero-D-manno-heptose. (na activatie wordt één van de OH groepen omgelegd door het epimerase RfaD) tal van decoraties ( versierselen ) : fosfo-ethanolamine, enz. verderop (outer core, O-antigen) : tal van bijzondere suikers: (dehydratatie, epimerisatie, reductie) GDP-mannose ==== GDP-L-fucose. TDP-glucose ==== TDP-L-rhamnose. CDP-glucose ==== CDP-abequose, CDP-paratose, CDP-tyvelose. vorming van deoxysuikers vanaf verschillende geactiveerde hexosen (deoxy, dideoxy) polymerisatie met transferasen die in de membraan zitten : een soort heen en weer drijven tot de geschikte donor gevonden wordt. (cfr. teichoïnezuursynthese)")
23
Determinantenketen (O-antigeen)
Opnieuw met bactoprenylfosfaat als drager. Eerst opbouw van monomeer-eenheid (tetrasaccharide) eerste stap : een fosfaatgroep gaat mee => pyrofosfaat (pyrofosfatase nodig voor recuperatie van bactoprenylfosfaat) Groeiende keten: aankoppeling aan het reducerende uiteinde van de groeiende herhaling. (bestaande keten wordt op nieuw monomeer geplaatst) Alle O-antigen polymerasen van verschillende organismen hebben minstens 12 TM. Lengte : (niet gelijk verdeeld: vooral lengtes 1 en 25 tot 35) de opbouw van de core zone gebeurt trapsgewijs: overzicht van de stappen in E.coli en Salmonella (bij Salmonella een amino-arabinose "decoratie) ( er zijn subtiele verschillen maar de basisstruktuur is vrij goed geconserveerd.)
 eerste stap : een fosfaatgroep gaat mee => pyrofosfaat (pyrofosfatase nodig. voor recuperatie van bactoprenylfosfaat) Groeiende keten: aankoppeling aan het reducerende uiteinde van de groeiende herhaling. (bestaande keten wordt op nieuw monomeer geplaatst) Alle O-antigen polymerasen van verschillende organismen hebben minstens 12 TM. Lengte : 1-40 (niet gelijk verdeeld: vooral lengtes 1 en 25 tot 35) de opbouw van de core zone gebeurt trapsgewijs: overzicht van de stappen in E.coli en Salmonella. (bij Salmonella een amino-arabinose decoratie) ( er zijn subtiele verschillen maar de basisstruktuur is vrij goed geconserveerd.)")
26
Teichoïnezuur, teichuronzuur, lipotechoïnezuur :
- Anionische polymeren. - Geven de cel een negatieve lading op de wand. Regelt daardoor mee de passage van ionen doorheen de celwand. - Bepalen tevens mee de poly-electrolyteigenschappen van de wand en zijn essentiëel voor de leefbaarheid en normale morfogenese. Voorkomen: - Alleen bij Gram-positieve bacteriën - lipoteichoïnezuren : steeds aanwezig, weinig beinvloed door de groeicondities - teichoïnezuur > < teichuronzuur: afhankelijk van groeicondities: indien fosfaatconcentratie laag wordt: switch van teichoïnezuur naar teichuronzuur Extractie: - peptidoglycaangebonden deel: onder licht zuur en licht alkalische voorwaarden (zie verder) - membraanverankerd deel: met hete 45% fenol-water Basisstructuur : - lichtvertakte polysaccharideketens aangekoppeld aan peptidoglycaan - zuur : fosfaatgroepen, teichuronzuur (=deels vervangen fosf. door carbonzure groep) (onder fosfaat limitatie)
 - membraanverankerd deel: met hete 45% fenol-water. Basisstructuur : - lichtvertakte polysaccharideketens aangekoppeld aan peptidoglycaan. - zuur : fosfaatgroepen, teichuronzuur (=deels vervangen fosf. door carbonzure groep) (onder fosfaat limitatie)")
28
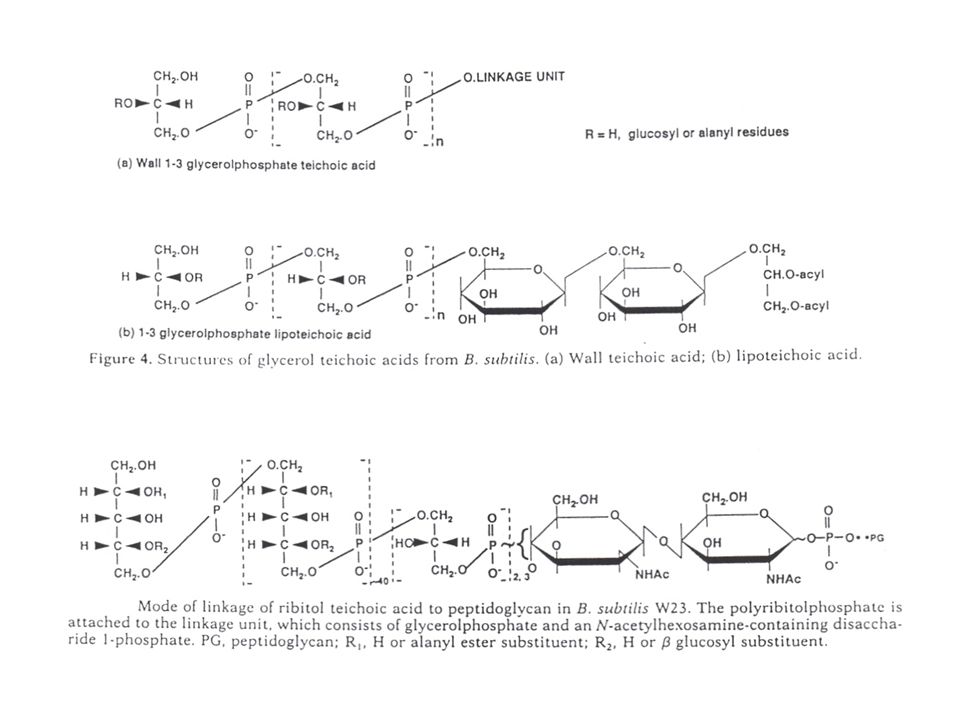
Keten (backbone) glycerolfosfaat (1,3) : soms (1,2), bij stammen van B. subtilis ribitolfosfaat (1,5) : bij S. aureus en diverse bacilli, waaronder ook stammen van B. subtilis; (maar ook andere varianten zijn gevonden: glucosylglycerolfosfaat, N-Ac-glucosamine-1-fosfaat -glycerolfosfaat, enz…) lengte van de ketens : ongeveer 40 eenheden (blijkbaar onafhankelijk van de groeicondities, dus regulatie gebeurt niet op dit niveau) vertakkingen: (R-groepen): op glycerol: a-glucosyl of D-alanylester (het D-ala-ester is zeer onstabiel) Als er een glycosylsubstituent is, kan ook dààrop een D-alanylester aanwezig zijn, die stabieler is en bijgevolg de analyse (van de andere positie) bemoeilijkt. op ribitol: op C2 : D-ala ester op C4 : b-glucosyl: van volledig tot nul naargelang de keten
 : bij S. aureus en diverse bacilli, waaronder ook stammen van B. subtilis; (maar ook andere varianten zijn gevonden: glucosylglycerolfosfaat, N-Ac-glucosamine-1-fosfaat -glycerolfosfaat, enz…) lengte van de ketens : ongeveer 40 eenheden (blijkbaar onafhankelijk van de groeicondities, dus regulatie gebeurt niet op dit niveau) vertakkingen: (R-groepen): op glycerol: a-glucosyl of D-alanylester (het D-ala-ester is zeer onstabiel) Als er een glycosylsubstituent is, kan ook dààrop een D-alanylester aanwezig zijn, die stabieler is en bijgevolg de analyse (van de andere positie) bemoeilijkt. op ribitol: op C2 : D-ala ester. op C4 : b-glucosyl: van volledig tot nul naargelang de keten.")
29
'Linkage unit' : B. subtilis W23 B. subtilis 168 op beide : 2 x glycerolfosfaat aan N-Ac-mannosamine – NAG –fosfaat; maar bij B.subtilis 168 ook N-Ac-galactosamine – glucose –fosfaat (10-30%, afhankelijk van groeicondities, bij 42°C zo goed als afwezig) glycerol – P – mannNAc – gluNAc – P- x=alkali-labiel x=zuurlabiel verklaart extractieresultaten (boven) Vrij constant teruggevonden bij alle andere bestudeerde species: blijkt goed geconserveerd. Enige variaties: in aantal glyc-P (2 of 3), glu i.p.v. NacMann-NH2, gluNH2 i.p.v. NAG
 glycerol – P – mannNAc – gluNAc – P- x=alkali-labiel. x=zuurlabiel. verklaart extractieresultaten (boven) Vrij constant teruggevonden bij alle andere bestudeerde species: blijkt goed geconserveerd. Enige variaties: in aantal glyc-P (2 of 3), glu i.p.v. NacMann-NH2, gluNH2 i.p.v. NAG.")
32
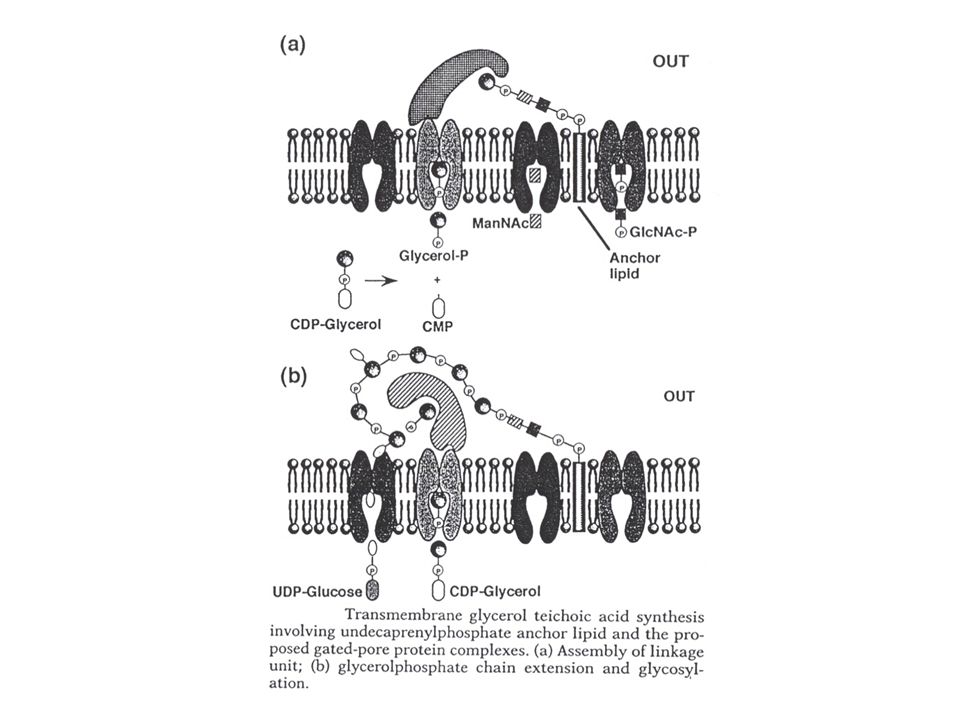
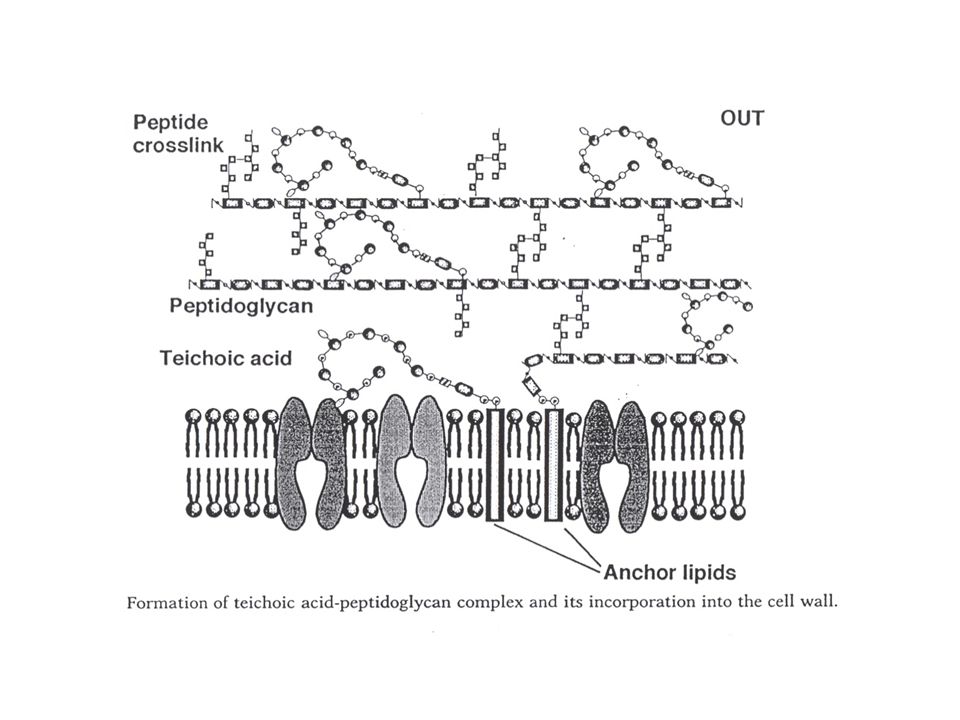
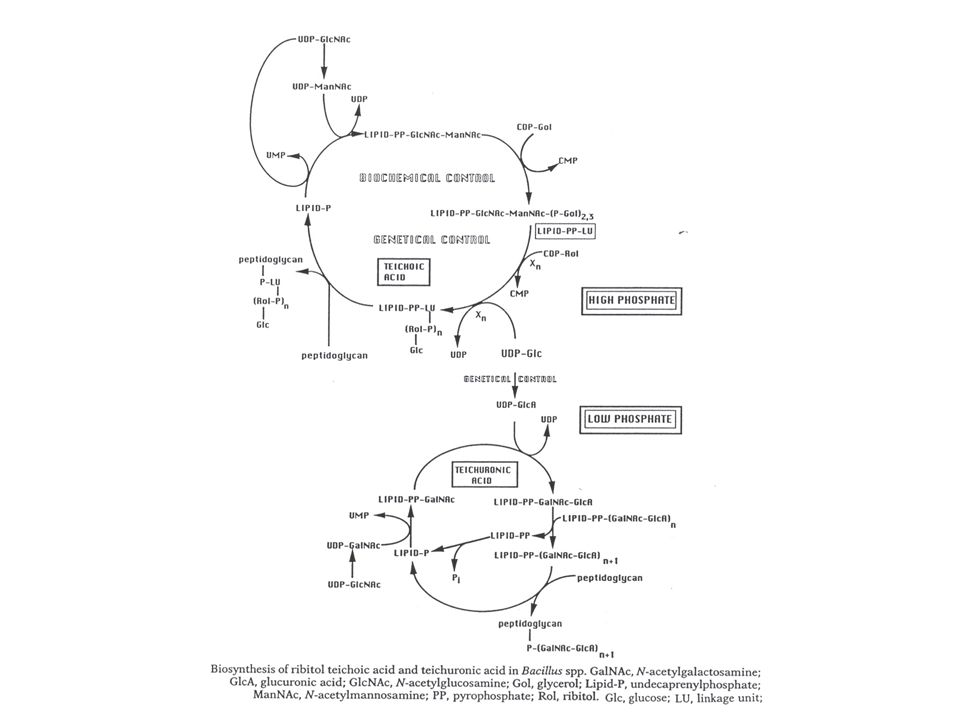
Opbouwmechanisme : teichoïnezuur
Verschillende mechanismen voorgesteld. Nogal wat hypothese (geen unificatie). CDP-glycerol en CDP-ribitol waren gevonden reeds lang voor de studie van de teichoïnezuren. Worden in het cytoplasma aangemaakt : cytidilyltransferasen (CDP-alditol pyrofosforylasen) : zijn gereguleerd door de fosfaatconcentratie: CDP-glyc : repressie van expressie EN inactivatie van bestaand enzyme (bij lage fosfaatconcentratie) CDP-ribitol : repressie van expressie, maar geen inactivatie van bestaand enzyme Eerste stap : N-Ac-Gluc aan bactoprenol, dan N-Ac-Mann : geactiveerd met UDP Dan : 2 of 3 glyc-P 's Dan : N eenheden ribitolfosfaat : aanhechting aan uiteinde van de keten, verst van het bactoprenyl verwijderd Modificatie zijketen: perifeer membraan glucosyltransferase (gtaA gen) (soms via bactoprenol) Afwerking cirkel : aanhechting aan peptidoglycaan: binnenin (maar “groeit naar buiten”, met de groei van het peptidoglycaan zelf). Geen pyrofosfatase nodig want bactoprenyl-P is het reactieproduct. Variant model : waarbij de ribitol(geglycosyleerd) apart op een lipidetransfercarrier wordt aangemaakt (zie Fig. hypotheses !!!)
. CDP-glycerol en CDP-ribitol waren gevonden reeds lang voor de studie van de teichoïnezuren. Worden in het cytoplasma aangemaakt : cytidilyltransferasen (CDP-alditol pyrofosforylasen) : zijn. gereguleerd door de fosfaatconcentratie: CDP-glyc : repressie van expressie EN inactivatie van bestaand enzyme (bij lage. fosfaatconcentratie) CDP-ribitol : repressie van expressie, maar geen inactivatie van bestaand enzyme. Eerste stap : N-Ac-Gluc aan bactoprenol, dan N-Ac-Mann : geactiveerd met UDP. Dan : 2 of 3 glyc-P s. Dan : N eenheden ribitolfosfaat : aanhechting aan uiteinde van de keten, verst van het bactoprenyl. verwijderd. Modificatie zijketen: perifeer membraan glucosyltransferase (gtaA gen) (soms via bactoprenol) Afwerking cirkel : aanhechting aan peptidoglycaan: binnenin (maar groeit naar buiten , met de groei. van het peptidoglycaan zelf). Geen pyrofosfatase nodig want bactoprenyl-P is het reactieproduct. Variant model : waarbij de ribitol(geglycosyleerd) apart op een lipidetransfercarrier wordt. aangemaakt (zie Fig. hypotheses !!!)")
35
Zijweg naar teichuronzuur: bij fosfaatlimitatie
Commentaar: - bij sommige bacteriestammen ook constitutief aanwezig naast teichoinezuur ( > < fosfaatgereguleerd) (o.a. sommige B. licheniformis stammen) - bij sommige stammen is het het enige (en constitutief) : bvb. B. megaterium en verschillende stammen van B. cereus (met N-Ac-mannosaminuronzuur) - bij B. subtilis W23 : (FIG) a, 1-4 glucuronzuur – a, 1-3 N-Ac-galactosamine Opbouw: - Geen linkage unit - Disaccharide aangebracht (Gal-N-Ac – Glucuronzuur) tussenschuiven in reeds langere keten die ook aan prenol hangt (=> prenol-pp : pyrofosfatase) - Na N aankoppelingen: aanhechting aan peptidoglycan : met fosfaat :=> dus prenol-p : geen pyrofosfatase nodig. Aanhechting aan C-6 van muraminezuur-eenheid. (Bij B. megaterium : trisaccharide repeat unit; bij Micrococcus luteus nog anders) HOE DOORHEEN DE MEMBRAAN ? Cfr. Peptidoglycaansynthese : flip-flop Hier tweede (geprefereerd) model : gebruik van pories : de feitelijke reacties gebeuren net buiten het cytoplasma. In ieder geval synthese in parallel met peptidoglycaan.
 (o.a. sommige B. licheniformis stammen) - bij sommige stammen is het het enige (en constitutief) : bvb. B. megaterium en. verschillende stammen van B. cereus (met N-Ac-mannosaminuronzuur) - bij B. subtilis W23 : (FIG) a, 1-4 glucuronzuur – a, 1-3 N-Ac-galactosamine. Opbouw: - Geen linkage unit. - Disaccharide aangebracht (Gal-N-Ac – Glucuronzuur) tussenschuiven in reeds. langere keten die ook aan prenol hangt (=> prenol-pp : pyrofosfatase) - Na N aankoppelingen: aanhechting aan peptidoglycan : met fosfaat :=> dus prenol-p : geen pyrofosfatase nodig. Aanhechting aan C-6 van muraminezuur-eenheid. (Bij B. megaterium : trisaccharide repeat unit; bij Micrococcus luteus nog anders) HOE DOORHEEN DE MEMBRAAN Cfr. Peptidoglycaansynthese : flip-flop. Hier tweede (geprefereerd) model : gebruik van pories : de feitelijke reacties gebeuren. net buiten het cytoplasma. In ieder geval synthese in parallel met peptidoglycaan.")
36
Opbouwmechanisme lipoteichoinezuur
Struktuur: FIG (zie boven) : in membraan : fosfaatester aan gentobiose (3-gentobiosyl-diglyceride) In membraan gehouden door intercalatie van de acylketens in het hydrofobe gebied van de membraan en gestabiliseerd door divalente kationenbruggen : - wordt losgemaakt door de-acylerend lipase - wordt losgemaakt door EDTA (Er kan wel een deel in de celwand “los” zitten, maar dit is iets anders dan hetgeen covalent covalent aan peptidoglycaan hangt) merk op : de configuratie van glycerol is anders (versus teichoinezuur) Opbouw: Geen bactoprenol nog: aanbreng rechtstreeks door glycolipide Fosfatidylglycerol => glycerolfosfaat naar glycolipide (lipide-gentobiose) + 1,2 diglyceride - seriele aanhechting van glyc-fosfaat op glyceroluiteinde van de groeiende keten - glycosylatie met tussenkomst van bactoprenylfosfaat (+ NAG, of galactose) - D-ala + ATP => D-alanyl-AMP, dan transfer op lipotech IN de membraan (recyclage van het 1,2 diglyceride door fosforylatie tot fosfatidinezuur en verder tot fosfatidylglycerol)
 : in membraan : fosfaatester aan gentobiose. (3-gentobiosyl-diglyceride) In membraan gehouden door intercalatie van de acylketens in het hydrofobe gebied van de. membraan en gestabiliseerd door divalente kationenbruggen : - wordt losgemaakt door de-acylerend lipase. - wordt losgemaakt door EDTA. (Er kan wel een deel in de celwand los zitten, maar dit is iets anders dan. hetgeen covalent covalent aan peptidoglycaan hangt) merk op : de configuratie van glycerol is anders (versus teichoinezuur) Opbouw: Geen bactoprenol nog: aanbreng rechtstreeks door glycolipide. Fosfatidylglycerol => glycerolfosfaat naar glycolipide (lipide-gentobiose) + 1,2 diglyceride. - seriele aanhechting van glyc-fosfaat op glyceroluiteinde van de groeiende keten. - glycosylatie met tussenkomst van bactoprenylfosfaat (+ NAG, of galactose) - D-ala + ATP => D-alanyl-AMP, dan transfer op lipotech IN de membraan. (recyclage van het 1,2 diglyceride door fosforylatie tot fosfatidinezuur en verder tot fosfatidylglycerol)")
37
Opbouw: Geen bactoprenol nog: aanbreng rechtstreeks door glycolipide Fosfatidylglycerol => glycerolfosfaat naar glycolipide (lipide-gentobiose) + 1,2 diglyceride - seriele aanhechting van glyc-fosfaat op glyceroluiteinde van de groeiende keten - glycosylatie met tussenkomst van bactoprenylfosfaat (+ NAG, of galactose) - D-ala + ATP => D-alanyl-AMP, dan transfer op lipotech IN de membraan (recyclage van het 1,2 diglyceride door fosforylatie tot fosfatidinezuur en verder tot fosfatidylglycerol) Lipoteichoinezuur kan bijdragen tot antigenische reacties. Steekt wat uit (ook andere teichoinezuren maar in mindere mate) => heeft een rol in interactie met dierlijke cellen. Teichoinezuren kunnen vrij in het medium terechtkomen, voor lipoteichoinezuren : vormen micellen. Interactie met andere componenten mogelijk: vb. virulentiegroep A streptococcen: hebben M-proteïne aan oppervlak - M-proteïne is positief geladen - bindt ionisch aan lipoteichoïnezuur, en exposeert => reactie met fibronectine van epitheliale cellen van mond en huid - functie: bescherming van de bacterie tegen fagocytose => veel antigenische variatie door mutaties in de M-proteïne sequentie.
 - D-ala + ATP => D-alanyl-AMP, dan transfer op lipotech IN de membraan. (recyclage van het 1,2 diglyceride door fosforylatie tot fosfatidinezuur en verder tot fosfatidylglycerol) Lipoteichoinezuur kan bijdragen tot antigenische reacties. Steekt wat uit (ook andere teichoinezuren. maar in mindere mate) => heeft een rol in interactie met dierlijke cellen. Teichoinezuren kunnen vrij in het medium terechtkomen, voor lipoteichoinezuren : vormen micellen. Interactie met andere componenten mogelijk: vb. virulentiegroep A streptococcen: hebben M-proteïne aan oppervlak. - M-proteïne is positief geladen. - bindt ionisch aan lipoteichoïnezuur, en exposeert. => reactie met fibronectine van epitheliale cellen van mond en huid. - functie: bescherming van de bacterie tegen fagocytose. => veel antigenische variatie door mutaties in de M-proteïne sequentie")
Verwante presentaties






>")













>")



