Download de presentatie
De presentatie wordt gedownload. Even geduld aub

1
Eilanden en evolutie Herpetologie 16 mei 2006

2
Overzicht Wat zijn eilanden Het belang in de biologie
Spectaculaire veranderingen fenotype Eilanden en evolutie Besluit

3
Wat zijn ‘eilanden’? Oceanische eilanden – vulkanische activiteit (Galapagos, Hawaii) Continentale eilanden – behoorden ooit tot het vasteland (UK) Maar ook andere geïsoleerde gebieden zoals: Landbouwgebieden Grotten Gefragmenteerde bossen Nunataks Vijvers, rivieren Gastheren (voor parasieten) Natuurreservaten …
 Natuurreservaten. …")
4
Eilanden en evolutie Biota op eilanden: veranderingen in fenotype
Oorzaken: Genetische divergentie? Founder effect Genetische drift Natuurlijke selectie Introgressie Fenotypische plasticiteit (omgeving)? Combinatie Eilandenreeks: herhaalde, discrete, relatief eenvoudige eenheden Dus ‘natuurlijke laboratoria’ (Whittaker, 1998) Testen algemene theorieën evolutie
 Combinatie. Eilandenreeks: herhaalde, discrete, relatief eenvoudige eenheden. Dus ‘natuurlijke laboratoria’ (Whittaker, 1998) Testen algemene theorieën evolutie.")
5
Eilanden en evolutie Eilanden kunnen dus een belangrijke rol spelen in evolutionaire studies: → Eerste naturalisten (Darwin, 1845; Wallace, 1859) → Hedendaagse evolutiebiologen (Losos, 2004) Populatiegenetica (variatie, populatiegenetische processen) Phylogenetica (evolutionaire relaties, vorm en functie) Ecologische perspectieven (invloed interacties waargenomen patroon, invasies exoten en hun impact, rol in volledige ecosysteem) Recente studies tonen zeer snelle evolutionaire veranderingen aan
 Populatiegenetica (variatie, populatiegenetische processen) Phylogenetica (evolutionaire relaties, vorm en functie) Ecologische perspectieven (invloed interacties waargenomen patroon, invasies exoten en hun impact, rol in volledige ecosysteem) Recente studies tonen zeer snelle evolutionaire veranderingen aan.")
6
Soortenrijkdom – oppervlakte relatie
Relatie tussen oppervlakte (A) van een eiland en de soortenrijkdom (S) Grote eilanden meer soorten Eilanden algemeen: disproportioneel hoge biodiversiteit & endemen i.v.m. vasteland S = Caz S = aantal soorten C = constante welke een maat is voor het aantal soorten per eenheid van oppervlakte A = oppervlakte Z = constante welke de helling is van de relatie tussen S en A
 van een eiland en de soortenrijkdom (S) Grote eilanden meer soorten. Eilanden algemeen: disproportioneel hoge biodiversiteit & endemen i.v.m. vasteland. S = Caz. S = aantal soorten. C = constante welke een maat is voor het aantal soorten per eenheid van oppervlakte. A = oppervlakte. Z = constante welke de helling is van de relatie tussen S en A.")
7
Soortenrijkdom – oppervlakte relatie
Andere factoren (soms inherent aan A) Heterogeniteit Topografisch reliëf Mate van verstoring Klimaat Aantal soorten beïnvloed door: Immigratie Locale extinctie (predatie, voedselschaarste) Emigratie (dispersie, migratie, …)
 Heterogeniteit. Topografisch reliëf. Mate van verstoring. Klimaat. Aantal soorten beïnvloed door: Immigratie. Locale extinctie (predatie, voedselschaarste) Emigratie (dispersie, migratie, …)")
8
Variatie in fenotype Morfologie, gedrag, ecologie, life history
Lichaamsgrootte Gigantisme Nanisme Herbivorie (afwezigheid predatie, voedseltekort) Tamheid (afwezigheid predatie) Vleugelloosheid (afwezigheid predatie) … → Eerste drie punten uitgewerkt
 Tamheid (afwezigheid predatie) Vleugelloosheid (afwezigheid predatie) … → Eerste drie punten uitgewerkt.")
9
Variatie in fenotype Morfologie, gedrag, ecologie, life history
Lichaamsgrootte Gigantisme Nanisme Herbivorie Tamheid Vleugelloosheid Tweehuizigheid …

10
Gigantisme Moa (Dinornis sp.) (Bunce et al., 2003)
3 Soorten beschreven 250 kg, 1 – 2 m schofthoogte Genetische studie: 1 soort Sexuele dimorfie vb. Ecological release door afwezigheid zoogdieren verlies vliegkracht (en energetische kost) afname metabolische snelheid
 afname metabolische snelheid.")
11
Gigantisme Moa (Dinornis sp.) (Bunce et al., 2003)
Discrepantie tss genetische homogeniteit & morfologische variatie!

12
Gigantisme Reuzenarend (Harpagornis moorei) van Nieuw-Zeeland (Bunce et al., 2005)
10 – 15 kg, VS: 2 – 3 m Zustertaxon: ‘dwerg’ → morfologische plasticiteit

13
Gigantisme Reuzenarend (Harpagornis moorei) van Nieuw-Zeeland (Bunce et al., 2005)
Potentiële snelheid verandering in grootte Opmerkelijke evolutionaire processen op eilanden

14
Variatie in fenotype Morfologie, gedrag, ecologie, life history
Lichaamsgrootte Gigantisme Nanisme Vb. Dwergolifanten Allopatrische speciatie Herbivorie Tamheid Vleugelloosheid …

15
Nanisme dwergolifanten
Verspreiding: Meditterrane eilanden Sardinië Sicilië & Malta Cyprus Cycladen Dodecanese eilanden Kanaaleilanden van Californië Wrangel eiland Indonesië Flores Tior Andere kleine Sunda eilanden Sulawesi Enkele duizenden jaren tijd: herhaalde evolutie tot dwergvormen Schofthoogte: 1.5 m!

16
Nanisme Amphibia Miniaturisatie: Brachycepahlidae Sooglossidae
(Psyllophryne didactyla, Brazilië) Leptodactylidae (Eleutherodactylus iberica, Cuba) Sooglossidae (Sooglossus gardineri, Seychellen) Microhylidae (Microhyla ornata, Borneo)
 Leptodactylidae. (Eleutherodactylus iberica, Cuba) Sooglossidae. (Sooglossus gardineri, Seychellen) Microhylidae. (Microhyla ornata, Borneo)")
17
Relaties binnen de groepen
Consistente trend naar: Gigantisme Nanisme binnen een bepaalde groep, vb.: Serpentes Tendens tot nanisme op eilanden (Case, 1978) Tendens tot gigantisme op eilanden (Forsman, 1991; Mori, 1994) Sommigen claimen gigantisme als een regel voor slangen (vb. Rodriguez & Drummond, 2000)
 Tendens tot gigantisme op eilanden (Forsman, 1991; Mori, 1994) Sommigen claimen gigantisme als een regel voor slangen (vb. Rodriguez & Drummond, 2000)")
18
Relaties binnen de groepen
Studie Boback (2002) Fylogenetisch geïnformeerde studie effect naar verandering in lichaamsgrootte bij Serpentes (Boidae, Colubridae, Elaphidae & Viperidae)
 Fylogenetisch geïnformeerde studie effect naar verandering in lichaamsgrootte bij Serpentes (Boidae, Colubridae, Elaphidae & Viperidae)")
19
Relaties binnen de groepen
Resultaten: Adders: afname in grootte ( 11 op 12) Colubers: geen patroon
 Colubers: geen patroon.")
20
Relaties binnen de groepen
Viperidae versus Colubridae Verklaringen: foerageerstrategie (Boback, 2003) Sit en wait (Viperidae) Actief (Colubridae) Broedende zeevogels: trend naar gigantisme Squamaten: trend naar nanisme
 Sit en wait (Viperidae) Actief (Colubridae) Broedende zeevogels: trend naar gigantisme. Squamaten: trend naar nanisme.")
21
Relaties binnen de groepen
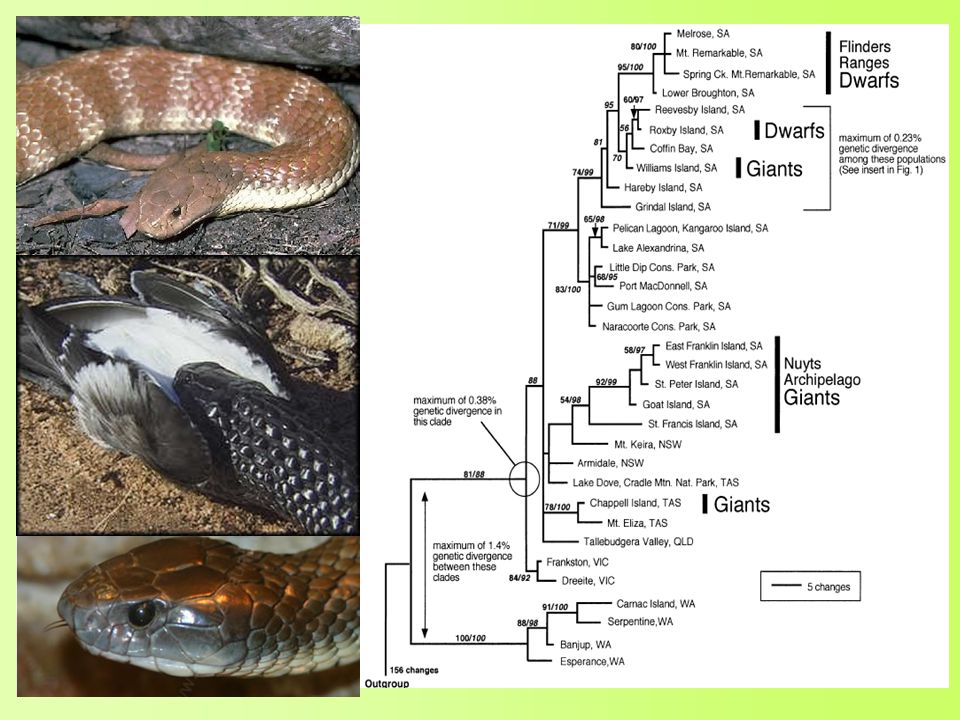
Trend binnen genera: Australische tijgerslangen (Notechis sp.) – Elaphidae (Keogh et al., 2005) Gigantisme en nanisme Nanisme: prooien kleiner dan vasteland Gigantisme: seizoenale prooibeschikbaarheid
 – Elaphidae (Keogh et al., 2005) Gigantisme en nanisme. Nanisme: prooien kleiner. dan vasteland. Gigantisme: seizoenale. prooibeschikbaarheid.")
23
Relaties binnen de groepen: Notechis sp.
Onafhankelijk ontstaan Sterke selectie ↔ beschikbare prooien Niet gezamenlijke evolutionaire afkomst Adaptieve plasticiteit Snelheid van evolutie (Haldanes) H = (dwerg) H = – (reus) Snelle evolutionaire verandering Voorbeeld herhaalde morfologische divergentie
 H = (dwerg) H = – (reus) Snelle evolutionaire verandering. Voorbeeld herhaalde morfologische divergentie.")
24
Is Testudo weissingeri een aparte evolutionaire lijn?
Afwijkende vormen niet enkel op klassieke eilanden: natuurlijke patchiness Testudo weissingeri: landschildpad die extreem droge gebieden bewoond ‘woestijnachtige’ eilandjes In 1996 beschreven als nieuwe Europese landschildpaddensoort

25
Is Testudo weissingeri een aparte evolutionaire lijn?
Klassiek 4 soorten Europese Testudo Fylogenetische relaties tussen Palearctisch genus Testudo Testudo weissingeri synoniem T. marginata (van der Kuyl et al., 2002; Fritz et al., 2005)
")
26
T. weissingeri parafyletisch genest binnen T. marginata
Subspecies T. graeca ondersteund Klassieke soorten binnen genus Testudo ondersteund

27
Is Testudo weissingeri een aparte evolutionaire lijn?
Kleine grootte Suboptimale omstandigheden (Fritz et al., 2005) Belang van kennis fylogenie! 4 soorten geaccepteerd 27 soorten (vb. Guyot – Jackson, 2004; Pieh & Perälä, 2004) ‘Grijze literatuur’ Exclusief morfologische kenmerken Belang rekenschap inter- en intraspecifieke diversiteit
 Belang van kennis fylogenie! 4 soorten geaccepteerd. 27 soorten (vb. Guyot – Jackson, 2004; Pieh & Perälä, 2004) ‘Grijze literatuur’ Exclusief morfologische kenmerken. Belang rekenschap inter- en intraspecifieke diversiteit.")
28
Is Testudo weissingeri een aparte evolutionaire lijn?
Diagnostische kenmerken T. weissingeri Kleine adulten (Fritz et al., 2005) Carapax – patronen (Artner, 1996)
 Carapax – patronen (Artner, 1996)")
29
Is de Komodo-varaan een dwerg?
Komodo – varaan: Archetype gigantisme Voedde zich met dwergolifanten Tijdens Pleistoceen: 4 keer grotere varanen leefden in Australië Sommige auteurs: Komodo – varaan = insulaire dwerg

30
Fenomenen verklaren Feiten gekend, oorzaken?
Welke factoren bevoordelen een soort om haar dimensies te veranderen? Wat bepaalt of een soort een reus of een dwerg wordt (t.o.v. vasteland vorm)?
")
31
Evolutie in lichaamsgrootte bij insulaire vertebraten: de eiland -regel
Trend naar gigantisme bij kleine soorten en nanisme bij grote soorten Bestaan er gerelateerde patronen van variatie in lichaamsgrootte (e.g. isolatie en oppervlakte)? Causale basis voor deze patronen?
 Causale basis voor deze patronen")
32
Evolutie in lichaamsgrootte bij insulaire vertebraten: de eiland -regel
Algemener dan aangenomen (Lomolino, 2005) Vogels: toename grootte bek bij insulaire populaties (Lack, 1947; Grant, 1965) Cassuaris Nieuw Guinea (Casuarius bennetti) klein ↔ Australië (Casuarius casuarinus) (Wallace, 1857)
 Vogels: toename grootte bek bij insulaire populaties (Lack, 1947; Grant, 1965) Cassuaris Nieuw Guinea (Casuarius bennetti) klein ↔ Australië (Casuarius casuarinus) (Wallace, 1857)")
33
Evolutie in lichaamsgrootte bij insulaire vertebraten: de eiland -regel
Reptielen: Diversiteit evolutie lichaamsgrootte (Foster, 1965) Gigantisme: meer algemeen bij leguanen, herbivore hagedissen, tijgerslangen en vermoedelijk ook landschildpadden
 Gigantisme: meer algemeen bij leguanen, herbivore hagedissen, tijgerslangen en vermoedelijk ook landschildpadden.")
34
Optimale grootte Soortspecifieke
Optimale lichaamsgrootte (bouwplan) (Case, 1978) Ecologische strategie (Noback & Guyer, 2003) Kleinere meer geïsoleerde, ecologisch eenvoudigere gemeenschappen: Kleine soorten: gigantisme Grote soorten: nanisme
 (Case, 1978) Ecologische strategie (Noback & Guyer, 2003) Kleinere meer geïsoleerde, ecologisch eenvoudigere gemeenschappen: Kleine soorten: gigantisme. Grote soorten: nanisme.")
35
Optimale grootte

36
Optimale grootte Gradiënt van verkleinende oppervlakte en afname van de soortendiversiteit leidt tot gemiddelde toename van lichaamsgrootte (Marquet & Taper, 1998) Lichaamsgrootte: invloed op Immigratie – potentieel Ecologische interacties Benodigde hulpbronnen Vogels: ecological release afwezigheid terrestrische zoogdieren
 Lichaamsgrootte: invloed op. Immigratie – potentieel. Ecologische interacties. Benodigde hulpbronnen. Vogels: ecological release afwezigheid terrestrische zoogdieren.")
37
Optimale grootte Vogels: toename in lichaamsgrootte (vb. Moa)
Shift dieet Bezetten niches typisch grote grazers Mate van verandering: afhankelijk organisme Kenmerken exploiteren hulpbronnen Interspecifieke interacties Immigratiekarakteristieken Kenmerken eilanden Primaire productiviteit / beschikbare hulpbronnen Diversiteit / intensiteit ecologische interacties Isolatie (leeftijd & geografische afstand)
")
38
Optimale grootte

39
Variatie in fenotype Morfologie, gedrag, ecologie, life history
Lichaamsgrootte Gigantisme Nanisme Herbivorie Tamheid Vleugelloosheid Tweehuizigheid …

40
Herbivorie Verschillende vormen herbivorie: Vegetatieve delen Fruit
Floraal voedsel: nectar, pollen Rol hagedissen traditioneel: Zeldzaam Onbelangrijk

41
Interactie hagedis & plant
2 Stadia reproductieve cycli mogelijke rol Pollinatie Dispersie zaden Pollinatie door dieren lang aanschouwt als exclusief domein van: Nectarivore invertebraten (insecten) Vogels (kolibri) Zoogdieren (Megachiroptera)
 Vogels (kolibri) Zoogdieren (Megachiroptera)")
42
Pollinatie: Podarcis lilfordi Voorbeeld van rechtstreekse pollinatie
Balearische endeem Bezoekt bloemen natieve Euphorbia dendroides Sterk geconcentreerde nectar Bij hoge densiteit: bezoek aan bloemen 8 keer hoger dan insecten P. lilfordi: belangrijke pollinator van minstens 23 planten (Traveset & Saez, 1997)
")
43
Pollinatie: Niveoscincus microlepidotus Voorbeeld van inrechtstreekse pollinatie
Kleine, abundante sneeuwskink (Tasmanië) Bezoekt Richea scoparia Scheurt calyptra (gefuseerde sepalen) open (omhult antheren en nectar): eet calyptra en nectar Blootstelling insecten: pollinatie
 Bezoekt Richea scoparia. Scheurt calyptra (gefuseerde sepalen) open (omhult antheren en nectar): eet calyptra en nectar. Blootstelling insecten: pollinatie.")
44
Interactie hagedis & plant
Zoöchorie (dier gemedieerde dispersie) lang aanschouwd als exclusief domein van: Vogels Zoogdieren Frugivorie relatief algemeen Oude groepen; belangrijke dispersers eerste gymnospermen en angiospermen Moderne reptielen: enkel landschildpadden (25%) Rol hagedissen belangrijk! (Borzi, 1911) Hagedissen Nieuw Zeeland (Whittaker, 1980)
 lang aanschouwd als exclusief domein van: Vogels. Zoogdieren. Frugivorie relatief algemeen. Oude groepen; belangrijke dispersers eerste gymnospermen en angiospermen. Moderne reptielen: enkel landschildpadden (25%) Rol hagedissen belangrijk! (Borzi, 1911) Hagedissen Nieuw Zeeland (Whittaker, 1980)")
45
Interactie hagedis & plant
Dus: hagedis als mutualistische factor genegeerd Fruitetend: 202 hagedissoorten in 19 families 68 % Eilandsoorten Aanschouwd als: ware carnivoor met slechts 1 % ware herbivoor

46
Interactie hagedis & plant
Herbivorie komt voor op vasteland, maar voornamelijk op eilanden Insulaire hagedissen drinken nectar, eten fruitpulp, ondanks lage hoeveelheid proteïne Waarom ? Densiteits compensatie Dieet expansie Lage predatiedruk Algemeen: geen coëvolutie (breed trofisch spectrum)
")
47
Oorzaken herbivorie Eilandfenomeen: waarom meer mutualistische hagedissen? 4 Potentiële redenen meer florale hulpbronnen en fruit in dieet : Surplus floraal voedsel & fruit Schaarste arthropoden Grotere hagedissen gepreadapteerd voor herbivoor dieet Gereduceerd predatierisico

48
1. Surplus floraal voedsel & fruit
Productie nectar / fruit niet toegenomen Minder geoogst Meeste pollinators: insecten Meeste frugivoren: vogels & zoogdieren Hebben lagere densiteit

49
2. Schaarste arthropoden
Klassieke nichetheorie: Lagere soortendensiteit (MacArthur & Wilson, 1967) Zwakkere interspecifieke competitie Niche - expansie Toename in densiteit Fenomeen: densiteits – compensatie Eilanden armer in soortenrijkdom insecten & abundantie (Janzen, 1973; Case, 1983) Biomassa arthropoden 4 keer lager Arthropoden doen niet aan densiteits - compensatie
 Zwakkere interspecifieke competitie. Niche - expansie. Toename in densiteit. Fenomeen: densiteits – compensatie. Eilanden armer in soortenrijkdom insecten & abundantie (Janzen, 1973; Case, 1983) Biomassa arthropoden 4 keer lager. Arthropoden doen niet aan densiteits - compensatie.")
50
3. Grotere hagedissen gepreadapteerd herbivoor dieet
Vegetatieve delen (bladeren en stengels) Veel cellulose Moeilijker te verteren dan pollen / nectar Het digestief systeem van herbivore hagedissen is dikwijls gespecialiseerd Voor digestieve & antipredator – redenen: Evolutie herbivorie gecorreleerd toename lichaamsgrootte Enkel hagedissen > 100 g succesvolle vegetariërs Vb. leguaan (uitz. varanen)
 Veel cellulose. Moeilijker te verteren dan pollen / nectar. Het digestief systeem van herbivore hagedissen is dikwijls gespecialiseerd. Voor digestieve & antipredator – redenen: Evolutie herbivorie gecorreleerd toename lichaamsgrootte. Enkel hagedissen > 100 g succesvolle vegetariërs. Vb. leguaan (uitz. varanen)")
51
3. Grotere hagedissen gepreadapteerd herbivoor dieet
Hagedissen < 100 g eten arthropoden Makkelijk verteerbaar voedsel (pollen & nectar): geen specifieke adaptaties spijsverteringsapparaat Kleine hagedissen op eilanden ≠ klassieke dichotomie (Pough) Kleine artropodeneters versus grote herbivoren
: geen specifieke adaptaties spijsverteringsapparaat. Kleine hagedissen op eilanden ≠ klassieke dichotomie (Pough) Kleine artropodeneters versus grote herbivoren.")
52
Voorbeelden afwijkingen dichotomie van Pough
Canarische lacertide hagedis: Gallotia galloti 55 g maar fruit belangrijkste component van het dieet Lepidophyma smithii (Midden – Amerika) 25 g Dieet: 91 % fruit
 25 g. Dieet: 91 % fruit.")
53
4. Gereduceerd predatierisico
Lager predatierisico (Szarski, 1962) Zoektijd Verteertijd Maar ook fruitetende hagedissen in predatorrijke habitats Vb. Platysaurus capensis Habitat extreem weinig arthropoden Beschikbaarheid belangrijker dan predatierisico (Olesen & Valido, 2003)
 Zoektijd. Verteertijd. Maar ook fruitetende hagedissen in predatorrijke habitats. Vb. Platysaurus capensis. Habitat extreem weinig arthropoden. Beschikbaarheid belangrijker dan predatierisico (Olesen & Valido, 2003)")
54
Variatie in fenotype Morfologie, gedrag, ecologie, life history
Lichaamsgrootte Gigantisme Nanisme Herbivorie Tamheid Vleugelloosheid Tweehuizigheid …

55
Tamheid Mogelijke oorzaak: verlies aan predatoren Grote vogels:
Gewicht derde macht! Vleugels gigantisch groot vleugelloos

56
Studie van evolutie Voorgaande voorbeelden spectaculair, maar veranderingen in fenotype meestal subtieler Systeem = monofyletische groep van soorten beperkt klein, geïsoleerd habitat Oceanische eilanden Hamvraag: welke evolutionaire processen speelden een rol in de divergentie die bestaat tussen eilandpopulaties?

57
Studie van evolutie Oorzaken: Genetische divergentie?
Founder effect Genetische drift Natuurlijke selectie Introgressie Fenotypische plasticiteit (omgeving)? Combinatie
 Combinatie.")
58
Oorzaak van divergentie: Genetische drift
Belangrijk lange duur (effectieve) populatiegrootte (Berry et al., 1992) Verlies / fixatie allelen Afname heterozygositeit Toename genetische populatiedifferentiatie
 populatiegrootte (Berry et al., 1992) Verlies / fixatie allelen. Afname heterozygositeit. Toename genetische populatiedifferentiatie.")
59
Oorzaak van divergentie: Genetische drift
Drift basis fenotypische divergentie Morfologie/prestatie en ‘fitness’ geen biologisch significant verband → Random walks

60
Oorzaak van divergentie: Natuurlijke selectie
- Differentiële selectie tgv verschillen abiotische en biotische condities

61
Oorzaak van divergentie: Hybridisatie/introgressie
Evolutie moleculaire technieken: komt meer voor dan aanvankelijk gedacht Soortvreemde allelen van sym- of parapatrische soorten Hagedissen Middelands zeegebied: Podarcis sicula & Podarcis tiliguerta (Capula, 2002) Podarcis sicula & Podarcis melisellensis (Gorman et al., 1975) Podarcis sicula & Podarcis raffonei (Capula, 1993) Podarcis sicula & Podarcis wagleriana (Capula, 1993)
 Podarcis sicula & Podarcis melisellensis (Gorman et al., 1975) Podarcis sicula & Podarcis raffonei (Capula, 1993) Podarcis sicula & Podarcis wagleriana (Capula, 1993)")
63
Evolutie op eilanden Anagenese (evolutionaire verandering binnen een lijn) is basis cladogenese Soort drukt relatie uit tot andere ‘individuen’ welke de nazaten zijn van dezelfde ‘ouders’

64
Besluit: evolutie op eilanden
Waargenomen spectaculaire fenotypes (vb. gigantisme) niet exclusief voor eilanden Voorkomen wel frequenter
 niet exclusief voor eilanden. Voorkomen wel frequenter.")
65
Besluit: evolutie op eilanden
Eilanden ideale systemen voor studie fundamenteel ecologische problemen Nicheverschuiving Ecological release Competitie Predatie Eilanden ‘natuurlijke laboratoria’ testen algemene theoriën evolutie

Verwante presentaties